Old World megadroughts and pluvials during the Common Era
1
2015
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
Cooling and societal change during the Late Antique Little Ice Age from 536 to around 660 AD
1
2016
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
2
1976
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 树木生长环境和自身遗传因素均能影响树轮密度与气候的关系,不同地形条件下决定树轮密度大小的主要环境因子可能不同,而遗传因素控制了树木应对环境变化的生理机制[3,5].此外,树轮密度的大小还可能受人为干扰、虫害、获取树轮密度方法的影响,掩盖了原本气候信号.因此,影响树轮密度与气候关系的要素和过程相对复杂. ...
神农架地区树轮δ 18O序列的气候指示意义
1
2018
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
神农架地区树轮δ 18O序列的气候指示意义
1
2018
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
大别山地区黄山松和油松树轮宽度的气候意义
2
2012
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 树木生长环境和自身遗传因素均能影响树轮密度与气候的关系,不同地形条件下决定树轮密度大小的主要环境因子可能不同,而遗传因素控制了树木应对环境变化的生理机制[3,5].此外,树轮密度的大小还可能受人为干扰、虫害、获取树轮密度方法的影响,掩盖了原本气候信号.因此,影响树轮密度与气候关系的要素和过程相对复杂. ...
大别山地区黄山松和油松树轮宽度的气候意义
2
2012
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 树木生长环境和自身遗传因素均能影响树轮密度与气候的关系,不同地形条件下决定树轮密度大小的主要环境因子可能不同,而遗传因素控制了树木应对环境变化的生理机制[3,5].此外,树轮密度的大小还可能受人为干扰、虫害、获取树轮密度方法的影响,掩盖了原本气候信号.因此,影响树轮密度与气候关系的要素和过程相对复杂. ...
全球落叶松属树轮气候学研究进展
3
2013
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 树轮密度的大小还可能受到人为干扰、虫害、树轮密度获取方法等的影响.Franceschini等[55]认为,间伐可使树木获取的光照和水分增加,抵抗环境胁迫的能力增强,其密度大小受气候变化的影响可能变小,但由于采样地生境较好,研究结果并未证实其假设.他指出需进一步在环境相对恶劣条件下采样来验证该假设.此外,树木因虫害而落叶,光合作用减弱,用于细胞径向生长和细胞壁增厚的光合产物减少,导致早材密度偏大而晚材密度偏小[56].然而在出现气候极端的年份树轮密度也会出现类似的情况,并且极端气候也易引发虫害,因此树轮密度、虫害、极端气候之间的关系较为复杂,目前主要是结合同一地区非宿主植物的树轮年表以及虫害记录来剔除虫害噪声[6].此外,近年相关研究表明,相比于X射线法,树轮径向解剖技术获得的参数能精确地反映密度变化,其获取的LWD与温度关系在高低频域上均较好[36,57]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
全球落叶松属树轮气候学研究进展
3
2013
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 树轮密度的大小还可能受到人为干扰、虫害、树轮密度获取方法等的影响.Franceschini等[55]认为,间伐可使树木获取的光照和水分增加,抵抗环境胁迫的能力增强,其密度大小受气候变化的影响可能变小,但由于采样地生境较好,研究结果并未证实其假设.他指出需进一步在环境相对恶劣条件下采样来验证该假设.此外,树木因虫害而落叶,光合作用减弱,用于细胞径向生长和细胞壁增厚的光合产物减少,导致早材密度偏大而晚材密度偏小[56].然而在出现气候极端的年份树轮密度也会出现类似的情况,并且极端气候也易引发虫害,因此树轮密度、虫害、极端气候之间的关系较为复杂,目前主要是结合同一地区非宿主植物的树轮年表以及虫害记录来剔除虫害噪声[6].此外,近年相关研究表明,相比于X射线法,树轮径向解剖技术获得的参数能精确地反映密度变化,其获取的LWD与温度关系在高低频域上均较好[36,57]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
中国利用树轮资料重建干湿变化研究进展
2
2016
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
中国利用树轮资料重建干湿变化研究进展
2
2016
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Annual increments of juniper dwarf shrubs above the tree line on the central Tibetan Plateau: A useful climatic proxy
1
2012
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
Tree ring density-based warm-season temperature reconstruction since A.D. 1610 in the eastern Tibetan Plateau
4
2015
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Cool North European summers and possible links to explosive volcanic eruptions
1
2013
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
Multivariate techniques for specifying tree-growth and climate relationships and for reconstructing anomalies in paleoclimate
1
1971
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
A 263-year record of summer temperature for northern Quebec reconstructed from tree-ring data and evidence of a major climatic shift in the early 1800's
1
1988
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
Radiodensitometric-dendroclimatological conifer chronologies from Lapland (Scandinavia) and the Alps (Switzerland)
1
1988
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
July-August temperature at Edinburgh between 1721 and 1975 from tree-ring density and width data
1
1984
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
Red spruce tree-ring widths and densities in Eastern North America as indicators of past climate
2
1986
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
Summer temperatures across northern North America: Regional reconstructions from 1760 using tree-ring densities
3
1994
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
树轮密度、稳定C同位素对过去近100 a陕西黄陵季节气温与降水的恢复
4
1997
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... [17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
树轮密度、稳定C同位素对过去近100 a陕西黄陵季节气温与降水的恢复
4
1997
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... [17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
用树轮密度及宽度资料重建新疆吉木萨尔县的季节降水和最高温度
3
1998
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
用树轮密度及宽度资料重建新疆吉木萨尔县的季节降水和最高温度
3
1998
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Post-industrial late summer warming recorded in tree-ring density in the eastern Tibetan Plateau
3
2020
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
Eastern Mediterranean summer temperatures since 730 CE from Mt. Smolikas tree-ring densities
1
2020
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
Concord and discord among Northern Hemisphere paleotemperature reconstructions from tree rings
2
2019
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... [21,22,23,24]. ...
Northern Hemisphere temperature anomalies during the 1450 s period of ambiguous volcanic forcing
2
2017
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 历史或地质时期火山喷发时间主要基于文献和冰芯记录数据来确定的,但有时候其定年存在误差[97].由于树轮MXD与温度相关性高,其能为火山喷发事件提供准确定年.如Schneider等[98]根据树轮MXD重建的北半球温度序列,发现过去千年有14个显著降温事件,并与冰芯中的火山喷发记录相吻合,为火山喷发提供了确切年代;Esper等[22]用20个MXD年表重建了北半球过去600 a温度变化序列,结果显示1453年降温程度最大,表明15世纪中期的火山喷发事件发生在1452年,而不是1458年;Briffa等[99]分析了北半球区域树轮密度年表,推测Billy Mitchell火山爆发可能在1495年或1587年. ...
August-September temperature variability on the Tibetan Plateau: Past, present and future
2
2019
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
Assessing non-linearity in European temperature-sensitive tree-ring data
2
2020
... 树木年轮具有分辨率高、定年准确、分布广泛等特点,是重建过去气候变化的重要代用资料[1,2,3],常用的代用指标有宽度、密度和同位素[4,5,6,7].树轮密度指木材的组织密度,包括早材最小密度(minimum earlywood density, MID)、早材平均密度(mean earlywood density, EWD)、晚材平均密度(mean latewood density, LWD)和晚材最大密度(maximum latewood density, MXD),其大小与早晚材细胞壁的厚度及细胞的大小有关.相对于宽度,树轮密度受生长季前期气候条件影响较小,能够更好地揭示生长季气候变化[8,9,10].20世纪60年代,国际上开启了基于树轮密度的气候重建工作[11].之后,MXD被广泛应用于区域气候重建[12,13,14,15,16].国内起步相对较晚,始于20世纪90年代[17,18].目前,国内外学者一方面力求利用密度年表重建更长时间尺度的气候序列[19,20,21];另一方面则是建立高分辨率的树轮密度网络,以此提高重建结果可信度,更好地揭示区域气候变化过程[21,22,23,24]. ...
... 综合树轮密度在其他方面的应用来看,树轮密度用于评估火山爆发的降温程度还存在争议,原因是树轮密度与温度关系受到了光照、伪轮等其他因子的影响,因此,在严格控制数据质量的同时,今后需关注光照对树轮密度与温度关系的影响.对于树轮密度研究中存在的分异性问题,多数解释是由于温度与树轮密度之间的相关性降低,与降水的相关性升高,因此树轮密度与气候变量的关系不一定是线性关系,今后可考虑用非线性模型解决该问题[24].此外,树轮密度与气候变量的关系并非一成不变,重建方程未必能代表所有时间段的树轮密度与气候因子的关系,因此今后的研究可比较不同校准方法重建同一个气候变量所产生的偏差[60]. ...
Scientific merits and analytical challenges of tree‐ring densitometry
2
2019
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... [25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
Cell size and wall dimensions drive distinct variability of earlywood and latewood density in Northern Hemisphere conifers
3
2017
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 国内外树轮密度重建工作主要分布于高纬度或高海拔地区,以重建生长季或生长季末期的温度为主,但重建的方差解释量存在空间和树种差异,而该差异可能源于环境、实验误差、复本量、标准化方法的影响.亚洲和北美洲的树轮密度年表多在1000 a以下,比欧洲短.重建工作涉及的树种包括云杉、落叶松、冷杉、松树、黄杉、铁杉等多个属的树种,但不同树种树轮密度揭示气候变化的能力可能存在差异,因此找出适合于重建区域气候变化的最优树种是今后树轮密度研究关注的重点之一,而对于利用多个树种密度重建空间大尺度的气候变化时,则需要考虑树种差异的影响.此外,国际上各研究区基本上都是以晚材密度重建生长季温度历史为主,但其实早材密度指标也包含有丰富的气候信息[26,32],今后重建工作可基于其他密度指标,或利用不同的数学方法将不同密度指标合成新的指标来开展,从而更好地揭示过去气候变化.在重建的气候变量方面,可以考虑其他要素,如海平面气压变化等. ...
Diverse climate sensitivity of Mediterranean tree-ring width and density
4
2010
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... [27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
Responses to climate by tree-ring widths and maximum latewood densities of two Abies species at upper and lower altitudinal distribution limits in central Japan
8
2011
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
28]
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
28]
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
28]
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
29]
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... 海拔对树轮密度与气候关系的影响主要体现在气候条件和土壤持水能力差异2个方面[32].其一,高海拔地区气候寒冷,其MXD的增长主要受生长季温度的控制[32],如Tatras、Norikura及长白山地区高海拔的MXD与生长季温度的相关明显高于低海拔[28,32,40].其二,不同海拔地区土壤持水能力的差异可能影响树轮密度对降水的响应,如Tatras低海拔山脉的土壤由石灰岩风化而成,储水能力差,树木易遭受干旱胁迫,MXD对7月干旱响应敏感;而高海拔地区土层的母质为花岗岩,土壤的蓄水能力较低海拔强,降水对MXD影响较弱[32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
... 由于遗传因素的影响,不同树种或同树种雌雄株应对环境变化的生理机制也有差异,从而影响树轮密度对气候变化的响应[41,45].树种差异会影响树轮密度对气候响应的敏感程度和时间.如地中海地区栎树(Quercus ilex)扎根深,在干旱条件下仍可以发生蒸发;而松树扎根浅,易因干旱引起气孔关闭,减弱蒸发.因此,栎树的树轮细胞更易丢失水分而发生变形,易发生密度突变,对气候变化更敏感[52].再如漠河兴安落叶松(Larix gmelinii)的晚材细胞加厚的活跃高峰可能比樟子松(Pinus sylvestris var. mongolica)早,其MXD对温度的最敏感时段比樟子松有所提前[41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
A Pinus cembra L. tree-ring record for late spring to late summer temperature in the Rhaetian Alps, Italy
1
2019
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
Different climate response of three tree ring proxies of Pinus sylvestris from the Eastern Carpathians, Romania
2
2019
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
Different maximum latewood density and blue intensity measurements techniques reveal similar results
2
2018
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
Climate sensitivity of high- and low-elevation Larix decidua MXD chronologies from the Tatra Mountains
9
2020
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
32]
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... 海拔对树轮密度与气候关系的影响主要体现在气候条件和土壤持水能力差异2个方面[32].其一,高海拔地区气候寒冷,其MXD的增长主要受生长季温度的控制[32],如Tatras、Norikura及长白山地区高海拔的MXD与生长季温度的相关明显高于低海拔[28,32,40].其二,不同海拔地区土壤持水能力的差异可能影响树轮密度对降水的响应,如Tatras低海拔山脉的土壤由石灰岩风化而成,储水能力差,树木易遭受干旱胁迫,MXD对7月干旱响应敏感;而高海拔地区土层的母质为花岗岩,土壤的蓄水能力较低海拔强,降水对MXD影响较弱[32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
... [32],如Tatras、Norikura及长白山地区高海拔的MXD与生长季温度的相关明显高于低海拔[28,32,40].其二,不同海拔地区土壤持水能力的差异可能影响树轮密度对降水的响应,如Tatras低海拔山脉的土壤由石灰岩风化而成,储水能力差,树木易遭受干旱胁迫,MXD对7月干旱响应敏感;而高海拔地区土层的母质为花岗岩,土壤的蓄水能力较低海拔强,降水对MXD影响较弱[32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
... ,32,40].其二,不同海拔地区土壤持水能力的差异可能影响树轮密度对降水的响应,如Tatras低海拔山脉的土壤由石灰岩风化而成,储水能力差,树木易遭受干旱胁迫,MXD对7月干旱响应敏感;而高海拔地区土层的母质为花岗岩,土壤的蓄水能力较低海拔强,降水对MXD影响较弱[32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
... [32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
... 综合国内外树轮密度响应研究工作,发现采样地多分布在高海拔或高纬地区,少数研究分布在干旱的山区.由于采样地地理位置、生长环境的差异,树轮密度对气候响应的关系错综复杂.在高大山体树木的生长上限及高纬地区树线北界,MXD能更好地揭示温度变化;而在干旱的地区,树轮密度序列中含有降水信号.然而目前鲜少有研究分析干热河谷地区的树轮密度对气候的响应,今后树轮密度研究可选取该区进行采样,由于该区气候高温低湿,有可能从其树轮密度年表中提取出降水信号[32].此外,树轮密度与气候的关系还受到树种、海拔、坡向、人类活动、虫害等因子的影响,但各方面研究的数量偏少,且目前主要是根据相关系数或方差解释量大小来判断不同树种或不同海拔树轮密度与气候变量关系的差异,而该差异有可能是由样本量、实验误差等造成的,因此今后还需更多研究工作进一步验证.另外,间伐对树轮密度的影响本质是树木对光照、养分竞争带来的影响,但要寻找合适的间伐采样地还相对困难,今后研究可从种群密度的角度来分析竞争对树轮密度与气候关系的影响[60]. ...
... 国内外树轮密度重建工作主要分布于高纬度或高海拔地区,以重建生长季或生长季末期的温度为主,但重建的方差解释量存在空间和树种差异,而该差异可能源于环境、实验误差、复本量、标准化方法的影响.亚洲和北美洲的树轮密度年表多在1000 a以下,比欧洲短.重建工作涉及的树种包括云杉、落叶松、冷杉、松树、黄杉、铁杉等多个属的树种,但不同树种树轮密度揭示气候变化的能力可能存在差异,因此找出适合于重建区域气候变化的最优树种是今后树轮密度研究关注的重点之一,而对于利用多个树种密度重建空间大尺度的气候变化时,则需要考虑树种差异的影响.此外,国际上各研究区基本上都是以晚材密度重建生长季温度历史为主,但其实早材密度指标也包含有丰富的气候信息[26,32],今后重建工作可基于其他密度指标,或利用不同的数学方法将不同密度指标合成新的指标来开展,从而更好地揭示过去气候变化.在重建的气候变量方面,可以考虑其他要素,如海平面气压变化等. ...
Siberian tree-ring and stable isotope proxies as indicators of temperature and moisture changes after major stratospheric volcanic eruptions
4
2019
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
33]
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
Summer temperature variations in the European Alps, A.D. 755-2004
2
2006
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
Tree-ring width and maximum latewood density at the North American tree line: Parameters of climatic-change
5
1992
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
35]
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
35]
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
35]
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
Dendroclimatic potential of dendroanatomy in temperature-sensitive Pinus sylvestris
3
2020
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... 树轮密度的大小还可能受到人为干扰、虫害、树轮密度获取方法等的影响.Franceschini等[55]认为,间伐可使树木获取的光照和水分增加,抵抗环境胁迫的能力增强,其密度大小受气候变化的影响可能变小,但由于采样地生境较好,研究结果并未证实其假设.他指出需进一步在环境相对恶劣条件下采样来验证该假设.此外,树木因虫害而落叶,光合作用减弱,用于细胞径向生长和细胞壁增厚的光合产物减少,导致早材密度偏大而晚材密度偏小[56].然而在出现气候极端的年份树轮密度也会出现类似的情况,并且极端气候也易引发虫害,因此树轮密度、虫害、极端气候之间的关系较为复杂,目前主要是结合同一地区非宿主植物的树轮年表以及虫害记录来剔除虫害噪声[6].此外,近年相关研究表明,相比于X射线法,树轮径向解剖技术获得的参数能精确地反映密度变化,其获取的LWD与温度关系在高低频域上均较好[36,57]. ...
Blue intensity in Pinus sylvestris tree-rings: Developing a new palaeoclimate proxy
2
2007
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
Climate signals in tree-ring width, density and δ 13C from larches in Eastern Siberia (Russia)
2
2008
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
Development of tree-ring maximum latewood density chronologies for the western Tien Shan Mountains, China: Influence of detrending: Method and climate response
4
2013
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
39]
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
39]
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
The altitudinal variability and temporal instability of the climate-tree-ring growth relationships for Changbai larch (Larix olgensis Henry) in the Changbai mountains area, Jilin, Northeastern China
6
2016
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
40]
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
40]
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 海拔对树轮密度与气候关系的影响主要体现在气候条件和土壤持水能力差异2个方面[32].其一,高海拔地区气候寒冷,其MXD的增长主要受生长季温度的控制[32],如Tatras、Norikura及长白山地区高海拔的MXD与生长季温度的相关明显高于低海拔[28,32,40].其二,不同海拔地区土壤持水能力的差异可能影响树轮密度对降水的响应,如Tatras低海拔山脉的土壤由石灰岩风化而成,储水能力差,树木易遭受干旱胁迫,MXD对7月干旱响应敏感;而高海拔地区土层的母质为花岗岩,土壤的蓄水能力较低海拔强,降水对MXD影响较弱[32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
黑龙江漠河兴安落叶松与樟子松树轮生长特性及其对气候的响应
7
2005
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
41]
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
41]
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
41]
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 由于遗传因素的影响,不同树种或同树种雌雄株应对环境变化的生理机制也有差异,从而影响树轮密度对气候变化的响应[41,45].树种差异会影响树轮密度对气候响应的敏感程度和时间.如地中海地区栎树(Quercus ilex)扎根深,在干旱条件下仍可以发生蒸发;而松树扎根浅,易因干旱引起气孔关闭,减弱蒸发.因此,栎树的树轮细胞更易丢失水分而发生变形,易发生密度突变,对气候变化更敏感[52].再如漠河兴安落叶松(Larix gmelinii)的晚材细胞加厚的活跃高峰可能比樟子松(Pinus sylvestris var. mongolica)早,其MXD对温度的最敏感时段比樟子松有所提前[41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
... [41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
黑龙江漠河兴安落叶松与樟子松树轮生长特性及其对气候的响应
7
2005
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
41]
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
41]
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
41]
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 由于遗传因素的影响,不同树种或同树种雌雄株应对环境变化的生理机制也有差异,从而影响树轮密度对气候变化的响应[41,45].树种差异会影响树轮密度对气候响应的敏感程度和时间.如地中海地区栎树(Quercus ilex)扎根深,在干旱条件下仍可以发生蒸发;而松树扎根浅,易因干旱引起气孔关闭,减弱蒸发.因此,栎树的树轮细胞更易丢失水分而发生变形,易发生密度突变,对气候变化更敏感[52].再如漠河兴安落叶松(Larix gmelinii)的晚材细胞加厚的活跃高峰可能比樟子松(Pinus sylvestris var. mongolica)早,其MXD对温度的最敏感时段比樟子松有所提前[41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
... [41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
高频光密度测量法在崆峒山树轮年表建立中的应用
7
2010
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
42]
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
42]
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
42]
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
高频光密度测量法在崆峒山树轮年表建立中的应用
7
2010
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
42]
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
42]
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
42]
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
巩乃斯河源树木年轮密度年表特征分析
5
2010
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
43]
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [43](表2). ...
巩乃斯河源树木年轮密度年表特征分析
5
2010
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
43]
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [43](表2). ...
树轮灰度与树轮密度的对比分析及其对气候要素的响应
2
2011
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
树轮灰度与树轮密度的对比分析及其对气候要素的响应
2
2011
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
小五台山青杨雌雄植株树轮生长特性及其对气候变化的响应差异
5
2014
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
45]
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 由于遗传因素的影响,不同树种或同树种雌雄株应对环境变化的生理机制也有差异,从而影响树轮密度对气候变化的响应[41,45].树种差异会影响树轮密度对气候响应的敏感程度和时间.如地中海地区栎树(Quercus ilex)扎根深,在干旱条件下仍可以发生蒸发;而松树扎根浅,易因干旱引起气孔关闭,减弱蒸发.因此,栎树的树轮细胞更易丢失水分而发生变形,易发生密度突变,对气候变化更敏感[52].再如漠河兴安落叶松(Larix gmelinii)的晚材细胞加厚的活跃高峰可能比樟子松(Pinus sylvestris var. mongolica)早,其MXD对温度的最敏感时段比樟子松有所提前[41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
... [45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
小五台山青杨雌雄植株树轮生长特性及其对气候变化的响应差异
5
2014
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [
45]
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 由于遗传因素的影响,不同树种或同树种雌雄株应对环境变化的生理机制也有差异,从而影响树轮密度对气候变化的响应[41,45].树种差异会影响树轮密度对气候响应的敏感程度和时间.如地中海地区栎树(Quercus ilex)扎根深,在干旱条件下仍可以发生蒸发;而松树扎根浅,易因干旱引起气孔关闭,减弱蒸发.因此,栎树的树轮细胞更易丢失水分而发生变形,易发生密度突变,对气候变化更敏感[52].再如漠河兴安落叶松(Larix gmelinii)的晚材细胞加厚的活跃高峰可能比樟子松(Pinus sylvestris var. mongolica)早,其MXD对温度的最敏感时段比樟子松有所提前[41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
... [45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
Can tree-ring density data reflect summer temperature extremes and associated circulation patterns over Fennoscandia?
1
2017
... 揭示树轮密度指标与气候要素关系是重建过去气候变化的前提.近几十年来,国内外学者开展了大量树轮密度对气候变化的响应研究,取得了丰硕的研究成果.树轮密度的大小取决于树轮细胞的大小及其细胞壁的厚度.生长季前期的形成层活动较强,细胞分裂和扩大,树木以径向生长为主;生长季后期形成层活动减弱,细胞中合成细胞壁的物质沉积,细胞壁加厚,因此树轮早材密度较小而晚材密度较大[25,26,27].生长季或生长季末期较高的温度能加强植物的光合作用,有利于细胞壁加厚,增加树轮晚材密度[25],尤其是在树木生长受温度限制的地区,树轮晚材密度对温度的响应更敏感.如在Pyrenees、Alps、Carpathians、Tatras、Altai、Norikura等高山树线地区[27,28,29,30,31,32,33,34](表1),以及在北美、西伯利亚北部等高纬树木生长北界地区,其MXD与生长季温度有较好的正相关关系[35,36,37,38].中国天山、长白山、崆峒山等山区的MXD也是对夏季温度响应最强,但相关性偏低[39,40,41,42,43,44,45](表2).此外,MXD在反映极端温度和相应的大气环流形势上也具有一定的潜力,如芬诺斯坎迪亚的34个MXD年表的第一主成分能反映1901—1978年夏季极端高温天数的变化,方差解释量为50%,并通过对高度场、风场的分析发现:2个序列的正极值年份,反气旋多分布于芬诺斯坎迪亚的中东部,常伴有高空急流的经向移动[46]. ...
Minimum wood density of Juniperus thurifera is a robust proxy of spring water availability in a continental Mediterranean climate
3
2014
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
Minimum wood density of conifers portrays changes in early season precipitation at dry and cold Eurasian regions
5
2017
... International studies on responses of tree-ring density to climate change
Tab.1 | 研究区 | 树种 | 拉丁名 | 指标 | 海拔/m | 气候变量 | 相关系数/时段 | 文献来源 |
| Pyrenees | 欧洲赤松
山赤松
欧洲冷杉 | Pinus sylvestris
Pinus uncinata
Abies alba | MXD | 2010 | T(5—9月) | 0.80/1952—2002年 | [27] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1600 | T(7、9月) | 0.30/1979—2005年 | [28] |
| Mount Norikura | 富士山冷杉 | Abies veitchii | MXD | 1900 | T(7—9月) | 0.35/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 1900 | T(7—9月) | 0.34/1979—2005年 | [28] |
| Mount Norikura | 大白叶冷杉 | Abies mariesii | MXD | 2400 | T(7—9月) | 0.44/1979—2005年 | [28] |
| Rhaetian Alps, Italy | 瑞士五针松 | Pinus cembra | MXD | 2075~2350 | T(5—9月) | 0.60/1901—2015年 | [29] |
| Eastern Carpathians | 欧洲赤松 | Pinus sylvestris | MXD | 750~1200 | Tmax(7月30日—9月24日) | 0.63/1961—2013年 | [30] |
| Tatras | 欧洲云杉 | Picea abies | MXD | 1480 | T(4—9月) | 0.62/1901—2004年 | [31] |
| Dolina Mengusovska | 欧洲落叶松 | Larix decidua | MXD | 1450 | T(5—6月) | 0.68/1951—2012年 | [32] |
| Village Vernar | 欧洲落叶松 | Larix decidua | MXD | 850 | P(7月) | 0.43/1951—2012年 | [32] |
| Altai | 西伯利亚落叶松 | Larix sibirica | MXD | 2380 | T(6—7月) | 0.57/1963—2000年 | [33] |
| Yakutia | 卡氏落叶松 | Larix cajanderi | MXD | 22 | T(6—7月) | 0.55/1950—2000年 | [33] |
| Alps | 欧洲落叶松 | Larix decidua | MXD | >1500 | T(6—9月) | 0.73/1911—2003年 | [34] |
| Franklin Mountains | 白云杉 | Picea glauca | MXD | 653 | T(5—8月) | 0.45/1944—1977年 | [35] |
| Hornby Cabin | 白云杉 | Picea glauca | MXD | 143 | T(5—8月) | 0.58/1944—1977年 | [35] |
| Cri Lake | 白云杉 | Picea glauca | MXD | 108 | T(5—9月) | 0.50/1944—1977年 | [35] |
| Coppermine | 白云杉 | Picea glauca | MXD | 200 | T(5—8月) | 0.48/1944—1977年 | [35] |
| North-eastern Finland | 欧洲赤松 | Pinus sylvestris | MXD | 200 | T(4—9月) | 0.68/1876—2013年 | [36] |
| Laanila | 欧洲赤松 | Pinus sylvestris | MXD | 220 | T(6—8月) | 0.61/1958—2002年 | [37] |
| Suntar Khayata | 卡氏落叶松 | Larix cajanderi | MXD | 900 | Tmin(6—8月) | 0.67/1929—2000年 | [38] |
| Iberian Peninsula | 西班牙刺柏 | Juniperus thurifera | MID | 530~1375 | P(4—6月) | -0.67/1951—2000年 | [47] |
| Southern Urals | 欧洲赤松 | Pinus sylvestris | MID | 740 | P(5月) | -0.58/1950—2002年 | [48] |
| Khangai | 西伯利亚落叶松 | Larix sibirica | MID | 1920 | P(6月) | -0.57/1950—2002年 | [48] |
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
48]
| Sierra de Gúdar | 欧洲黑松 | Pinus nigra | MID | 1090 | P(5月) | -0.65/1950—2002年 | [48] |
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... [
48]
注:T表示平均温度,Tmax表示平均最高温度,Tmin表示平均最低温度;P表示降水.下同. ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... ,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
云杉树轮生长密度对气候要素的响应分析
3
2012
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
云杉树轮生长密度对气候要素的响应分析
3
2012
... Studies on responses of tree-ring density to climate change in China
Tab.2 | 样点 | 树种 | 海拔/m | 指标 | 响应变量 | 相关系数/时段 | 文献来源 |
| 吉普克 | 雪岭云杉 | 2555 | MXD | T(8月) | 0.35/1959—2004年 | [39] |
| 阿乌里亚乔克山 | 雪岭云杉 | 2690 | MXD | T(4—8月) | 0.42/1961—2005年 | [39] |
| 小白代沟 | 雪岭云杉 | 2682 | MXD | T(6—8月) | 0.33/1956—2006年 | [39] |
| 长白山 | 长白落叶松 | 1848 | MXD | Tmax(6—8月) | 0.38/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 1585 | MXD | Tmax(4—9月) | 0.44/1958—2008年 | [40] |
| 长白山 | 长白落叶松 | 983 | MXD | Tmin(8月) | -0.26/1958—2008年 | [40] |
| 漠河 | 兴安落叶松 | 650 | MXD | Tmax(7月) | 0.45/1922—1998年 | [41] |
| 漠河 | 兴安落叶松 | 650 | MID | Tmin(3月) | -0.323/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MXD | Tmax(8月) | 0.49/1922—1998年 | [41] |
| 漠河 | 樟子松 | 650 | MID | P(4月) | 0.343/1922—1998年 | [41] |
| 崆峒山 | 油松 | 1950 | MID | P(6—8月) | -0.4/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | EWD | P(6—8月) | -0.36/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | MXD | T(6—8月) | -0.26/1951—2006年 | [42] |
| 崆峒山 | 油松 | 1950 | LWD | T(6—9月) | -0.28/1951—2006年 | [42] |
| 艾肯达坂 | 天山云杉 | 2455 | MXD | T(7—8月) | 0.30/1968—2005年 | [43] |
| 巩乃斯林场北 | 天山云杉 | 1982 | EWD | P(6—7月) | -0.50/1968—2005年 | [43] |
| 艾肯达坂 | 雪岭云杉 | 2450 | MXD | Tmax(5—8月) | 0.304/1958—2008年 | [44] |
| 小五台山 | 青杨(雌) | 1600 | MXD | Tmax(8月) | 0.348/1982—2011年 | [45] |
| 小五台山 | 青杨(雄) | 1600 | MXD | Tmax(4月) | -0.429/1982—2011年 | [45] |
| 祁连山中段 | 青海云杉 | 3140~3390 | EWD | P(5月) | -0.623/1956—2009年 | [49] |
在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... 在树木生长受温度限制的地区,早材密度对温度的响应不同于晚材密度,温度的升高更有利于早材细胞的径向增大,而细胞壁加厚是次要的,因此高温导致早材密度更小[26].但干旱地区的树轮早材密度对生长季早期的降水更为敏感,当春季干旱稍明显时,早材细胞径向扩张减弱,引起密度增大[47,48];当干旱较严重时,早材细胞脱水,原生质与细胞壁收缩,在细胞壁上形成很多锐利的折叠,撕破原生质结构,导致早材密度增大[42-43,49].如Iberian Peninsula、Southern Urals Khangai、Sierra de Gúdar等气候干冷地区的MID与5—6月的降水呈显著的负相关关系[47,48](表1).中国崆峒山的早材密度与6—8月的降水呈负相关关系[42],祁连山中段的早材密度与5月降水负相关[49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
... [49],巩乃斯林场北的早材密度与6—7月的降水负相关[43](表2). ...
Effects of recent minimum temperature and water deficit increases on Pinus pinaster radial growth and wood density in Southern Portugal
2
2016
... 此外,树轮密度年内变化受到气候季节变化的影响.地中海地区生长季早期的干旱使树轮密度突然增大,秋季的多雨使晚材密度突然变小,从而形成伪轮[50,51,52].但由于早材树轮细胞的生长要消耗植物体内存储的养料,即早材密度还受到上年冬季气候的影响,所以早材中出现伪轮的情况较少,只有在极端干旱、寒冷的年份下才出现.而晚材密度主要受当年生长季气候的影响,所以出现变异的频次较多[50]. ...
... [50]. ...
Site conditions influence the climate signal of intra-annual density fluctuations in tree rings of Q-ilex L
2
2018
... 此外,树轮密度年内变化受到气候季节变化的影响.地中海地区生长季早期的干旱使树轮密度突然增大,秋季的多雨使晚材密度突然变小,从而形成伪轮[50,51,52].但由于早材树轮细胞的生长要消耗植物体内存储的养料,即早材密度还受到上年冬季气候的影响,所以早材中出现伪轮的情况较少,只有在极端干旱、寒冷的年份下才出现.而晚材密度主要受当年生长季气候的影响,所以出现变异的频次较多[50]. ...
... 坡向影响树轮密度对季节气候变化的敏感程度,如在地中海地区,南坡光周期更长,蒸发量更大,水分条件相对较差,其树轮密度更易因水分条件的变化而产生变异,因此南坡树轮比北坡树轮更易形成伪轮[51]. ...
Contrasting physiological responses to Mediterranean climate variability are revealed by intra-annual density fluctuations in tree rings of Quercus ilex L. and Pinus pinea L
2
2018
... 此外,树轮密度年内变化受到气候季节变化的影响.地中海地区生长季早期的干旱使树轮密度突然增大,秋季的多雨使晚材密度突然变小,从而形成伪轮[50,51,52].但由于早材树轮细胞的生长要消耗植物体内存储的养料,即早材密度还受到上年冬季气候的影响,所以早材中出现伪轮的情况较少,只有在极端干旱、寒冷的年份下才出现.而晚材密度主要受当年生长季气候的影响,所以出现变异的频次较多[50]. ...
... 由于遗传因素的影响,不同树种或同树种雌雄株应对环境变化的生理机制也有差异,从而影响树轮密度对气候变化的响应[41,45].树种差异会影响树轮密度对气候响应的敏感程度和时间.如地中海地区栎树(Quercus ilex)扎根深,在干旱条件下仍可以发生蒸发;而松树扎根浅,易因干旱引起气孔关闭,减弱蒸发.因此,栎树的树轮细胞更易丢失水分而发生变形,易发生密度突变,对气候变化更敏感[52].再如漠河兴安落叶松(Larix gmelinii)的晚材细胞加厚的活跃高峰可能比樟子松(Pinus sylvestris var. mongolica)早,其MXD对温度的最敏感时段比樟子松有所提前[41].但也存在同一地区不同树种树轮密度与气候关系基本一致的情况,如日本Norikura山1900 m处富士山冷杉与大白叶冷杉,其MXD对气候的响应基本一致[28](表1).因此,树种差异对树轮密度与气候关系的影响存在区域差异,仍需更多研究来验证.树木性别可能影响树轮密度对气候响应的时间,如小五台山青杨(Populus cathayana)雄株MXD对初春温度有负的响应,而雌株没有,其原因是合适的低温下,雌株萌发先于雄株,雄株在养分竞争中处于劣势,使得生长季节后期的形成层活动减弱,径向生长减缓,MXD增大[45].但目前只见青杨雌雄株树轮密度对气候响应差异的报道,其他树种雌雄异株的影响尚未可知. ...
The influence of elevational differences in absolute maximum density values on regional climate reconstructions
1
2015
... 海拔对树轮密度与气候关系的影响主要体现在气候条件和土壤持水能力差异2个方面[32].其一,高海拔地区气候寒冷,其MXD的增长主要受生长季温度的控制[32],如Tatras、Norikura及长白山地区高海拔的MXD与生长季温度的相关明显高于低海拔[28,32,40].其二,不同海拔地区土壤持水能力的差异可能影响树轮密度对降水的响应,如Tatras低海拔山脉的土壤由石灰岩风化而成,储水能力差,树木易遭受干旱胁迫,MXD对7月干旱响应敏感;而高海拔地区土层的母质为花岗岩,土壤的蓄水能力较低海拔强,降水对MXD影响较弱[32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
years of warm-season temperature variability in central Scandinavia inferred from tree-ring density
3
2016
... 海拔对树轮密度与气候关系的影响主要体现在气候条件和土壤持水能力差异2个方面[32].其一,高海拔地区气候寒冷,其MXD的增长主要受生长季温度的控制[32],如Tatras、Norikura及长白山地区高海拔的MXD与生长季温度的相关明显高于低海拔[28,32,40].其二,不同海拔地区土壤持水能力的差异可能影响树轮密度对降水的响应,如Tatras低海拔山脉的土壤由石灰岩风化而成,储水能力差,树木易遭受干旱胁迫,MXD对7月干旱响应敏感;而高海拔地区土层的母质为花岗岩,土壤的蓄水能力较低海拔强,降水对MXD影响较弱[32].此外,不同海拔MXD大小不同,并且在去除非气候因素影响时并不能完全消除此差异,如果不同海拔的样本量在时间上分布不均匀,会扭曲树轮密度年表的趋势,干扰气候信息的获取[53,54]. ...
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
Effect of thinning on the relationship between mean ring density and climate in black spruce (Picea mariana (Mill.) B.S.P.)
1
2018
... 树轮密度的大小还可能受到人为干扰、虫害、树轮密度获取方法等的影响.Franceschini等[55]认为,间伐可使树木获取的光照和水分增加,抵抗环境胁迫的能力增强,其密度大小受气候变化的影响可能变小,但由于采样地生境较好,研究结果并未证实其假设.他指出需进一步在环境相对恶劣条件下采样来验证该假设.此外,树木因虫害而落叶,光合作用减弱,用于细胞径向生长和细胞壁增厚的光合产物减少,导致早材密度偏大而晚材密度偏小[56].然而在出现气候极端的年份树轮密度也会出现类似的情况,并且极端气候也易引发虫害,因此树轮密度、虫害、极端气候之间的关系较为复杂,目前主要是结合同一地区非宿主植物的树轮年表以及虫害记录来剔除虫害噪声[6].此外,近年相关研究表明,相比于X射线法,树轮径向解剖技术获得的参数能精确地反映密度变化,其获取的LWD与温度关系在高低频域上均较好[36,57]. ...
Tree ring width and wood density as the indicators of climatic factors and insect outbreaks affecting spruce growth
1
2012
... 树轮密度的大小还可能受到人为干扰、虫害、树轮密度获取方法等的影响.Franceschini等[55]认为,间伐可使树木获取的光照和水分增加,抵抗环境胁迫的能力增强,其密度大小受气候变化的影响可能变小,但由于采样地生境较好,研究结果并未证实其假设.他指出需进一步在环境相对恶劣条件下采样来验证该假设.此外,树木因虫害而落叶,光合作用减弱,用于细胞径向生长和细胞壁增厚的光合产物减少,导致早材密度偏大而晚材密度偏小[56].然而在出现气候极端的年份树轮密度也会出现类似的情况,并且极端气候也易引发虫害,因此树轮密度、虫害、极端气候之间的关系较为复杂,目前主要是结合同一地区非宿主植物的树轮年表以及虫害记录来剔除虫害噪声[6].此外,近年相关研究表明,相比于X射线法,树轮径向解剖技术获得的参数能精确地反映密度变化,其获取的LWD与温度关系在高低频域上均较好[36,57]. ...
Were medieval warm-season temperatures in J?mtland, central Scandinavian Mountains, lower than previously estimated?
5
2019
... 树轮密度的大小还可能受到人为干扰、虫害、树轮密度获取方法等的影响.Franceschini等[55]认为,间伐可使树木获取的光照和水分增加,抵抗环境胁迫的能力增强,其密度大小受气候变化的影响可能变小,但由于采样地生境较好,研究结果并未证实其假设.他指出需进一步在环境相对恶劣条件下采样来验证该假设.此外,树木因虫害而落叶,光合作用减弱,用于细胞径向生长和细胞壁增厚的光合产物减少,导致早材密度偏大而晚材密度偏小[56].然而在出现气候极端的年份树轮密度也会出现类似的情况,并且极端气候也易引发虫害,因此树轮密度、虫害、极端气候之间的关系较为复杂,目前主要是结合同一地区非宿主植物的树轮年表以及虫害记录来剔除虫害噪声[6].此外,近年相关研究表明,相比于X射线法,树轮径向解剖技术获得的参数能精确地反映密度变化,其获取的LWD与温度关系在高低频域上均较好[36,57]. ...
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... [57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 就年表长度与重建序列长度来看,欧洲多千年长度的MXD年表,因Scandinavia地区的欧洲赤松活树的年龄可达500 a以上,死树年龄可达1000 a以上并且保存得较为完整[57],因此目前该区拥有世界上最长的MXD年表,长达2226 a(216 BC—AD 2010),由瑞典北部Torneträsk及Fennoscandia北部2条序列合成,并重建了过去2018 a(8 BC—AD 2010)温度变化[60].北美洲地区在21世纪10年代也将MXD年表延长到了千年尺度,目前最长MXD年表及重建序列为930 a(AD 1073—2002),其研究区位于Alaska的Firth河流域[69].中亚和日本地区目前的树轮密度年表与重建序列主要在三四百年间[74,75,76],相对偏短. ...
Tornetr?sk tree-ring width and density AD 500-2004: A test of climatic sensitivity and a new 1500-year reconstruction of north Fennoscandian summers
3
2008
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... “分异性”问题是指基于树轮代用资料重建的温度序列在近几十年没有显著上升的趋势,与器测记录产生了显著差异[69,109].最早在阿拉斯加地区白云杉树轮宽度与密度对气候响应的研究中发现“分异性”问题[67].随后,在白令海峡与楚科奇地区[66]、芬诺斯坎迪亚北部[58]、比利牛斯山脉[63]等高纬度地区的树轮密度研究中也出现了“分异性”问题.中低纬度地区出现“分异性”问题的研究相对较少.在中国青藏高原东南部也存在该问题,Li等[79]发现在2005年后MXD与温度相关性降低. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
Fennoscandia revisited: A spatially improved tree-ring reconstruction of summer temperatures for the last 900 years
3
2015
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 除用树轮密度单一指标重建外,还有研究认为MXD与EWD均受春季气候影响,两者存在一定相关性[71],为剔除生长季前期气候影响,利用MXD与EWD差值重建夏季平均温度变化过程,方差解释量高达70%(1855—2006年)[59].此外,也有研究尝试重建温度以外的其他气候变量.如Briffa等[72]用北半球341个MXD序列重建了海平面气压变化,但由于校准和验证时段过短、大气环流与树轮密度是间接关系等因素影响,重建结果并不理想.他指出,通过利用更长时间的器测温度序列,以及对降水敏感的代用资料,或许能改善重建结果的信度. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
Testing long-term summer temperature reconstruction based on maximum density chronologies obtained by reanalysis of tree-ring data sets from northernmost Sweden and Finland
5
2014
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 就年表长度与重建序列长度来看,欧洲多千年长度的MXD年表,因Scandinavia地区的欧洲赤松活树的年龄可达500 a以上,死树年龄可达1000 a以上并且保存得较为完整[57],因此目前该区拥有世界上最长的MXD年表,长达2226 a(216 BC—AD 2010),由瑞典北部Torneträsk及Fennoscandia北部2条序列合成,并重建了过去2018 a(8 BC—AD 2010)温度变化[60].北美洲地区在21世纪10年代也将MXD年表延长到了千年尺度,目前最长MXD年表及重建序列为930 a(AD 1073—2002),其研究区位于Alaska的Firth河流域[69].中亚和日本地区目前的树轮密度年表与重建序列主要在三四百年间[74,75,76],相对偏短. ...
... 综合国内外树轮密度响应研究工作,发现采样地多分布在高海拔或高纬地区,少数研究分布在干旱的山区.由于采样地地理位置、生长环境的差异,树轮密度对气候响应的关系错综复杂.在高大山体树木的生长上限及高纬地区树线北界,MXD能更好地揭示温度变化;而在干旱的地区,树轮密度序列中含有降水信号.然而目前鲜少有研究分析干热河谷地区的树轮密度对气候的响应,今后树轮密度研究可选取该区进行采样,由于该区气候高温低湿,有可能从其树轮密度年表中提取出降水信号[32].此外,树轮密度与气候的关系还受到树种、海拔、坡向、人类活动、虫害等因子的影响,但各方面研究的数量偏少,且目前主要是根据相关系数或方差解释量大小来判断不同树种或不同海拔树轮密度与气候变量关系的差异,而该差异有可能是由样本量、实验误差等造成的,因此今后还需更多研究工作进一步验证.另外,间伐对树轮密度的影响本质是树木对光照、养分竞争带来的影响,但要寻找合适的间伐采样地还相对困难,今后研究可从种群密度的角度来分析竞争对树轮密度与气候关系的影响[60]. ...
... 综合树轮密度在其他方面的应用来看,树轮密度用于评估火山爆发的降温程度还存在争议,原因是树轮密度与温度关系受到了光照、伪轮等其他因子的影响,因此,在严格控制数据质量的同时,今后需关注光照对树轮密度与温度关系的影响.对于树轮密度研究中存在的分异性问题,多数解释是由于温度与树轮密度之间的相关性降低,与降水的相关性升高,因此树轮密度与气候变量的关系不一定是线性关系,今后可考虑用非线性模型解决该问题[24].此外,树轮密度与气候变量的关系并非一成不变,重建方程未必能代表所有时间段的树轮密度与气候因子的关系,因此今后的研究可比较不同校准方法重建同一个气候变量所产生的偏差[60]. ...
Long-term summer temperature variations in the Pyrenees
3
2008
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... [61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
Estimating 750 years of temperature variations and uncertainties in the Pyrenees by tree-ring reconstructions and climate simulations
3
2012
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... [62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
New tree-ring evidence from the Pyrenees reveals Western Mediterranean climate variability since Medieval Times
5
2017
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... ,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... “分异性”问题是指基于树轮代用资料重建的温度序列在近几十年没有显著上升的趋势,与器测记录产生了显著差异[69,109].最早在阿拉斯加地区白云杉树轮宽度与密度对气候响应的研究中发现“分异性”问题[67].随后,在白令海峡与楚科奇地区[66]、芬诺斯坎迪亚北部[58]、比利牛斯山脉[63]等高纬度地区的树轮密度研究中也出现了“分异性”问题.中低纬度地区出现“分异性”问题的研究相对较少.在中国青藏高原东南部也存在该问题,Li等[79]发现在2005年后MXD与温度相关性降低. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
A 1200 + year reconstruction of temperature extremes for the northeastern Mediterranean region
2
2019
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
Spatial reconstruction of Scottish summer temperatures from tree rings
2
2017
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
Reconstructed warm season temperatures for Nome, Seward Peninsula, Alaska
3
2004
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... “分异性”问题是指基于树轮代用资料重建的温度序列在近几十年没有显著上升的趋势,与器测记录产生了显著差异[69,109].最早在阿拉斯加地区白云杉树轮宽度与密度对气候响应的研究中发现“分异性”问题[67].随后,在白令海峡与楚科奇地区[66]、芬诺斯坎迪亚北部[58]、比利牛斯山脉[63]等高纬度地区的树轮密度研究中也出现了“分异性”问题.中低纬度地区出现“分异性”问题的研究相对较少.在中国青藏高原东南部也存在该问题,Li等[79]发现在2005年后MXD与温度相关性降低. ...
Tree-ring width and density evidence of climatic and potential forest change in Alaska
3
1995
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... “分异性”问题是指基于树轮代用资料重建的温度序列在近几十年没有显著上升的趋势,与器测记录产生了显著差异[69,109].最早在阿拉斯加地区白云杉树轮宽度与密度对气候响应的研究中发现“分异性”问题[67].随后,在白令海峡与楚科奇地区[66]、芬诺斯坎迪亚北部[58]、比利牛斯山脉[63]等高纬度地区的树轮密度研究中也出现了“分异性”问题.中低纬度地区出现“分异性”问题的研究相对较少.在中国青藏高原东南部也存在该问题,Li等[79]发现在2005年后MXD与温度相关性降低. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
Temperature changes along the gulf of Alaska and the Pacific Northwest coast modeled from coastal tree rings
3
1996
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... [68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
Tree-ring-reconstructed summer temperatures from Northwestern North America during the last nine centuries
4
2013
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 就年表长度与重建序列长度来看,欧洲多千年长度的MXD年表,因Scandinavia地区的欧洲赤松活树的年龄可达500 a以上,死树年龄可达1000 a以上并且保存得较为完整[57],因此目前该区拥有世界上最长的MXD年表,长达2226 a(216 BC—AD 2010),由瑞典北部Torneträsk及Fennoscandia北部2条序列合成,并重建了过去2018 a(8 BC—AD 2010)温度变化[60].北美洲地区在21世纪10年代也将MXD年表延长到了千年尺度,目前最长MXD年表及重建序列为930 a(AD 1073—2002),其研究区位于Alaska的Firth河流域[69].中亚和日本地区目前的树轮密度年表与重建序列主要在三四百年间[74,75,76],相对偏短. ...
... “分异性”问题是指基于树轮代用资料重建的温度序列在近几十年没有显著上升的趋势,与器测记录产生了显著差异[69,109].最早在阿拉斯加地区白云杉树轮宽度与密度对气候响应的研究中发现“分异性”问题[67].随后,在白令海峡与楚科奇地区[66]、芬诺斯坎迪亚北部[58]、比利牛斯山脉[63]等高纬度地区的树轮密度研究中也出现了“分异性”问题.中低纬度地区出现“分异性”问题的研究相对较少.在中国青藏高原东南部也存在该问题,Li等[79]发现在2005年后MXD与温度相关性降低. ...
Wood density provides new opportunities for reconstructing past temperature variability from southeastern Australian trees
3
2016
... 国际上的重建工作多分布在Scandinavia[57,58,59,60]、Pyrenees[61,62,63]、Pindus[64]、Scotland[65]、Alaska[66,67,68,69]等高山高纬地区,且基本上是以MXD来重建区域生长季(4—9月)或生长季晚期(7—9月)的温度历史,其在器测时期温度重建的方差解释量均超过了50%,表明这些地区的MXD能较好地反映研究区温度变化.而在美洲西部、澳大利亚东南部等地区生长季或夏季温度重建的方差解释量相对较低,如Washington地区1905—1983年生长季温度的重建方差解释量为39%[68],Tasmania中部地区1960—2009年1月最高温度重建的方差解释量为34%[70],这可能是由于树轮密度对温度变化敏感度降低所引起,也有可能是复本量较少、标准化方法不同等因素造成的.如Pyrenees中部地区复本量由216个增至414个,重建的方差解释量由28.09%(1944—2005年)增至51.84%(1950—2014年)[61,63].虽然在Scandinavia地区研究中温度重建的方差解释量较高,但在AD 850—950年和AD 1100—1300年期间的复本量较低,导致重建不确定性较高.Linderholm等[57]在Zhang等[54]研究的基础上,增加了45个来自当地的干木材样本,删除了来自古建筑的样本,所重建的中世纪暖期的温度要比之前重建的低,但由于中世纪暖期的复本量还是偏少,目前该时期的温度变化还是难以确定.Dorado Liñán等[62]指出,当研究区地理范围较广时,直接使用所有测量序列建立区域标准化年表时,如没有满足区域标准化方法中复本量和年龄分类的要求,结果的可靠性可能会较差;而如果先建立不同样点的小区域年表、再将小区域年表合成大区域年表,则可能提高方差解释量. ...
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Blue intensity and density from northern Fennoscandian tree rings, exploring the potential to improve summer temperature reconstructions with earlywood information
1
2014
... 除用树轮密度单一指标重建外,还有研究认为MXD与EWD均受春季气候影响,两者存在一定相关性[71],为剔除生长季前期气候影响,利用MXD与EWD差值重建夏季平均温度变化过程,方差解释量高达70%(1855—2006年)[59].此外,也有研究尝试重建温度以外的其他气候变量.如Briffa等[72]用北半球341个MXD序列重建了海平面气压变化,但由于校准和验证时段过短、大气环流与树轮密度是间接关系等因素影响,重建结果并不理想.他指出,通过利用更长时间的器测温度序列,以及对降水敏感的代用资料,或许能改善重建结果的信度. ...
Tree-ring width and density data around the Northern Hemisphere: Part 2, spatio-temporal variability and associated climate patterns
1
2002
... 除用树轮密度单一指标重建外,还有研究认为MXD与EWD均受春季气候影响,两者存在一定相关性[71],为剔除生长季前期气候影响,利用MXD与EWD差值重建夏季平均温度变化过程,方差解释量高达70%(1855—2006年)[59].此外,也有研究尝试重建温度以外的其他气候变量.如Briffa等[72]用北半球341个MXD序列重建了海平面气压变化,但由于校准和验证时段过短、大气环流与树轮密度是间接关系等因素影响,重建结果并不理想.他指出,通过利用更长时间的器测温度序列,以及对降水敏感的代用资料,或许能改善重建结果的信度. ...
Boreal temperature variability inferred from maximum latewood density and tree-ring width data, Wrangell Mountain region, Alaska
2
2003
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
树轮最大密度记录的吉尔吉斯斯坦天山山区公元1650年以来的7—8月温度变化
2
2014
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 就年表长度与重建序列长度来看,欧洲多千年长度的MXD年表,因Scandinavia地区的欧洲赤松活树的年龄可达500 a以上,死树年龄可达1000 a以上并且保存得较为完整[57],因此目前该区拥有世界上最长的MXD年表,长达2226 a(216 BC—AD 2010),由瑞典北部Torneträsk及Fennoscandia北部2条序列合成,并重建了过去2018 a(8 BC—AD 2010)温度变化[60].北美洲地区在21世纪10年代也将MXD年表延长到了千年尺度,目前最长MXD年表及重建序列为930 a(AD 1073—2002),其研究区位于Alaska的Firth河流域[69].中亚和日本地区目前的树轮密度年表与重建序列主要在三四百年间[74,75,76],相对偏短. ...
树轮最大密度记录的吉尔吉斯斯坦天山山区公元1650年以来的7—8月温度变化
2
2014
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 就年表长度与重建序列长度来看,欧洲多千年长度的MXD年表,因Scandinavia地区的欧洲赤松活树的年龄可达500 a以上,死树年龄可达1000 a以上并且保存得较为完整[57],因此目前该区拥有世界上最长的MXD年表,长达2226 a(216 BC—AD 2010),由瑞典北部Torneträsk及Fennoscandia北部2条序列合成,并重建了过去2018 a(8 BC—AD 2010)温度变化[60].北美洲地区在21世纪10年代也将MXD年表延长到了千年尺度,目前最长MXD年表及重建序列为930 a(AD 1073—2002),其研究区位于Alaska的Firth河流域[69].中亚和日本地区目前的树轮密度年表与重建序列主要在三四百年间[74,75,76],相对偏短. ...
Tree ring density-based summer temperature reconstruction for Zajsan Lake area, East Kazakhstan
2
2012
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 就年表长度与重建序列长度来看,欧洲多千年长度的MXD年表,因Scandinavia地区的欧洲赤松活树的年龄可达500 a以上,死树年龄可达1000 a以上并且保存得较为完整[57],因此目前该区拥有世界上最长的MXD年表,长达2226 a(216 BC—AD 2010),由瑞典北部Torneträsk及Fennoscandia北部2条序列合成,并重建了过去2018 a(8 BC—AD 2010)温度变化[60].北美洲地区在21世纪10年代也将MXD年表延长到了千年尺度,目前最长MXD年表及重建序列为930 a(AD 1073—2002),其研究区位于Alaska的Firth河流域[69].中亚和日本地区目前的树轮密度年表与重建序列主要在三四百年间[74,75,76],相对偏短. ...
Warm-season annual to decadal temperature variability for Hokkaido, Japan, inferred from maximum latewood density (AD 1557-1990) and ring width data (AD 1532-1990)
2
2002
... 大多数针叶树种树轮密度均可用来进行气候重建(图1),但由于环境差异,导致重建所用树种也存在区域差异.欧洲Scandinavia山脉主要使用欧洲赤松[54,57,59-60,65],Pyrenees山脉采用山赤松(Pinus uncinata)[61,63],Pindus山脉采用波斯尼亚松(Pinus heldreichii)[64].美洲Alaska地区采用白云杉[66,69,73],Maine地区采用红云杉(Picea rubens)[15],中亚地区主要为雪岭云杉(Picea schrenkiana)[74]和西伯利亚落叶松[75],日本中部为库页云杉(Picea glehnii)[76].当进行大空间尺度重建时,研究者多利用不同树种的树轮密度资料,如Briffa等[16]采用云杉、落叶松、冷杉的MXD重建了北美北部地区夏季温度变化;Wiles等[68]用白云杉、北美云杉(Picea sitchensis)、花旗松(Pseudotsuga menziesii)和大果铁杉(Tsuga mertensiana)的MXD重建了阿拉斯加和太平洋西北部海岸4—9月的温度变化. ...
... 就年表长度与重建序列长度来看,欧洲多千年长度的MXD年表,因Scandinavia地区的欧洲赤松活树的年龄可达500 a以上,死树年龄可达1000 a以上并且保存得较为完整[57],因此目前该区拥有世界上最长的MXD年表,长达2226 a(216 BC—AD 2010),由瑞典北部Torneträsk及Fennoscandia北部2条序列合成,并重建了过去2018 a(8 BC—AD 2010)温度变化[60].北美洲地区在21世纪10年代也将MXD年表延长到了千年尺度,目前最长MXD年表及重建序列为930 a(AD 1073—2002),其研究区位于Alaska的Firth河流域[69].中亚和日本地区目前的树轮密度年表与重建序列主要在三四百年间[74,75,76],相对偏短. ...
Late summer temperature reconstruction based on tree-ring density for Sygera Mountain, southeastern Tibetan Plateau
3
2018
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
A 382-year reconstruction of August mean minimum temperature from tree-ring maximum latewood density on the southeastern Tibetan Plateau, China
2
2016
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Temperature reconstruction and volcanic eruption signal from tree-ring width and maximum latewood density over the past 304 years in the southeastern Tibetan Plateau
5
2017
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
... “分异性”问题是指基于树轮代用资料重建的温度序列在近几十年没有显著上升的趋势,与器测记录产生了显著差异[69,109].最早在阿拉斯加地区白云杉树轮宽度与密度对气候响应的研究中发现“分异性”问题[67].随后,在白令海峡与楚科奇地区[66]、芬诺斯坎迪亚北部[58]、比利牛斯山脉[63]等高纬度地区的树轮密度研究中也出现了“分异性”问题.中低纬度地区出现“分异性”问题的研究相对较少.在中国青藏高原东南部也存在该问题,Li等[79]发现在2005年后MXD与温度相关性降低. ...
Tree-ring density inferred late summer temperature variability over the past three centuries in the Gaoligong Mountains, southeastern Tibetan Plateau
1
2015
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
A 352-year record of summer temperature reconstruction in the western Tianshan Mountains, China, as deduced from tree-ring density
2
2013
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
树轮记录的贺兰山北部近208年5—7月温度变化
2
2011
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
树轮记录的贺兰山北部近208年5—7月温度变化
2
2011
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Reconstructing mean maximum temperatures of May-August from tree-ring maximum density in North Da Hinggan Mountains, China
3
2012
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
利用树轮密度重建新疆北部5—8月温度变化
4
2017
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... [84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
利用树轮密度重建新疆北部5—8月温度变化
4
2017
... 中国重建工作多是基于几个样点树轮密度资料开展的区域气候重建,分布在西南、西北和东北地区(图2),其中西南主要分布在青藏高原东南缘[77,78]、横断山脉及周边地区[9,79-80];西北在天山[81]、祁连山[70]、贺兰山[82]等地;东北在长白山[40]及大兴安岭[83].近几年来大空间尺度的重建也在开展,如Duan等[23]用了青藏高原地区17个采样点的树轮密度资料重建研究区自1572年来8—9月的温度变化,陈峰等[84]用新疆北部地区5个序列重建了研究区近353 a的5—8月温度变化.方差解释量同样存在空间差异(图2a),西南地区重建的结果相对较好,进一步验证了气候高寒区域的树轮晚材密度能够更好地揭示其生长季温度的历史变化. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... [84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
Absence of late-summer warming trend over the past two and half centuries on the eastern Tibetan Plateau
2
2014
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
May-June maximum temperature reconstruction from mean earlywood density in north Central China and its linkages to the summer monsoon activities
2
2014
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
以树木年轮密度资料重建鄂尔多斯中部地区6至10月降水量的变化
2
2013
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
以树木年轮密度资料重建鄂尔多斯中部地区6至10月降水量的变化
2
2013
... 重建气候要素存在空间差异(图2b).东北和西南地区夏季降水充沛,树木生长的主要限制因子为温度,多数工作采用MXD重建区域生长季温度[9,77,79,83,85].而西北地区气候干旱,温度和降水均有可能限制树木生长[17],使用的密度指标与重建要素相对复杂,如石门山低海拔地区生长季早期气候暖干,温度升高加剧土壤水分的蒸发,树轮密度受温度影响,故其EWD被用于重建5—6月的平均最高温度变化[86];吉木萨尔、陕西黄陵生长季早期的树轮生长主要受降水影响,因而其MID被用于重建生长季降水变化[17,18];鄂尔多斯地处中国北方干旱半干旱地区,其树轮晚材形成受到夏末到冬初的降水的影响,降水增多使树木生长季延长,树轮晚材多形成内径小、壁质厚的分生细胞,使得树轮晚材密度偏大,因此其MXD被用于重建6—10月的降水变化[87]. ...
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Temperature reconstruction from tree-ring maximum density of Balfour spruce in eastern Tibet, China
1
2010
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
A 449 year warm season temperature reconstruction in the southeastern Tibetan Plateau and its relation to solar activity
2
2014
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
采用高山松最大密度重建川西高原近百年夏季气温
1
2005
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
采用高山松最大密度重建川西高原近百年夏季气温
1
2005
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
树轮最大密度记录的贡嘎山区公元1837年以来的温度变化
1
2010
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
树轮最大密度记录的贡嘎山区公元1837年以来的温度变化
1
2010
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
Tree ring density-based summer temperature reconstruction for the central Hengduan Mountains in Southern China
1
2009
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
用天山雪岭云杉年轮最大密度重建新疆伊犁地区春夏季平均最高温度变化
1
2009
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
用天山雪岭云杉年轮最大密度重建新疆伊犁地区春夏季平均最高温度变化
1
2009
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
树轮记录的过去384a乌鲁木齐河源7月温度变化
2
2011
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
树轮记录的过去384a乌鲁木齐河源7月温度变化
2
2011
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
用西伯利亚落叶松年轮最大密度重建和布克赛尔5—8月份平均最高温度
1
2010
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
用西伯利亚落叶松年轮最大密度重建和布克赛尔5—8月份平均最高温度
1
2010
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
长白落叶松树轮生长对气候变化的响应以及对夏季气温的重建
2
2012
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
长白落叶松树轮生长对气候变化的响应以及对夏季气温的重建
2
2012
... 国内用于重建的树种也较多.中国西南地区针叶树种资源丰富,先后有研究使用川西云杉(Picea likiangensis)[19,78,85,88-89]、紫果云杉(Picea purpurea)[9]、高山松(Pinus densata)[90]及峨眉冷杉(Abies fabri)[91]、急尖长苞冷杉(Abies georgei)[77]、大果云杉(Picea asperata)[79]、麦吊云杉(Picea brachytyla)[92]等树种MXD进行气候重建.但目前该研究区各树种重建的方差解释量存在差异,如峨眉冷杉和高山松的重建方差解释量在50%以上,而急尖长苞冷杉和怒江红杉的方差解释量只有39%.该差异可能来源于遗传因素、环境或实验误差的影响,也可能是由于树轮密度揭示气候变化的能力存在种间差异,今后还需采取同一地点不同树种的树轮密度资料进行验证.西北和东北地区用于重建的树种相对偏少,但均为各研究区内树轮生长对气候响应较为敏感的树种[6-7,84]:天山地区为雪岭云杉[18,81,93-94]、阿尔泰山为西伯利亚落叶松[84,95]、祁连山地区青海云杉(Picea crassifolia)[70],石门山、贺兰山、陕西黄陵等地为油松(Pinus tabuliformis)[17,82,86-87],东北地区主要是兴安落叶松[83]、长白落叶松(Larix olgensis)[96](图2c). ...
... 相对于欧美地区,国内密度年表与重建的温度序列相对较短.在西南地区,青藏高原东部马尔康的树轮密度年表最长,为640 a (AD 1375—2014)[19];基于MXD重建的最长温度序列为449 a (AD 1563—2011)[89](图2d),来源于青藏高原东南部.西北地区阿尔泰山的树轮密度最长年表为521 a (AD 1488—2008)[84],但重建的最长温度序列来源于乌鲁木齐河源地区,为384 a (AD 1623—2006)[94].东北长白山密度年表最长为393 a (AD 1615—2008),其重建序列为238 a (AD 1762—1999)[96].由此可见,树轮密度年表最长的地方,其重建序列不一定最长,这表明需要更多年龄更大的样芯,通过增加更早历史时期的复本量来延长重建长度. ...
Tree-ring evidence of the widespread effects of explosive volcanic-eruptions
2
1995
... 历史或地质时期火山喷发时间主要基于文献和冰芯记录数据来确定的,但有时候其定年存在误差[97].由于树轮MXD与温度相关性高,其能为火山喷发事件提供准确定年.如Schneider等[98]根据树轮MXD重建的北半球温度序列,发现过去千年有14个显著降温事件,并与冰芯中的火山喷发记录相吻合,为火山喷发提供了确切年代;Esper等[22]用20个MXD年表重建了北半球过去600 a温度变化序列,结果显示1453年降温程度最大,表明15世纪中期的火山喷发事件发生在1452年,而不是1458年;Briffa等[99]分析了北半球区域树轮密度年表,推测Billy Mitchell火山爆发可能在1495年或1587年. ...
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
Corrigendum: A new archive of large volcanic events over the past millennium derived from reconstructed summer temperatures
1
2017
... 历史或地质时期火山喷发时间主要基于文献和冰芯记录数据来确定的,但有时候其定年存在误差[97].由于树轮MXD与温度相关性高,其能为火山喷发事件提供准确定年.如Schneider等[98]根据树轮MXD重建的北半球温度序列,发现过去千年有14个显著降温事件,并与冰芯中的火山喷发记录相吻合,为火山喷发提供了确切年代;Esper等[22]用20个MXD年表重建了北半球过去600 a温度变化序列,结果显示1453年降温程度最大,表明15世纪中期的火山喷发事件发生在1452年,而不是1458年;Briffa等[99]分析了北半球区域树轮密度年表,推测Billy Mitchell火山爆发可能在1495年或1587年. ...
Influence of volcanic eruptions on Northern Hemisphere summer temperature over the past 600 years
1
1998
... 历史或地质时期火山喷发时间主要基于文献和冰芯记录数据来确定的,但有时候其定年存在误差[97].由于树轮MXD与温度相关性高,其能为火山喷发事件提供准确定年.如Schneider等[98]根据树轮MXD重建的北半球温度序列,发现过去千年有14个显著降温事件,并与冰芯中的火山喷发记录相吻合,为火山喷发提供了确切年代;Esper等[22]用20个MXD年表重建了北半球过去600 a温度变化序列,结果显示1453年降温程度最大,表明15世纪中期的火山喷发事件发生在1452年,而不是1458年;Briffa等[99]分析了北半球区域树轮密度年表,推测Billy Mitchell火山爆发可能在1495年或1587年. ...
基于树轮资料初探过去千年强火山喷发与青藏高原东部温度变化关系
1
2016
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
基于树轮资料初探过去千年强火山喷发与青藏高原东部温度变化关系
1
2016
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
Signals and memory in tree-ring width and density data
1
2015
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
Journal of Climate
1
1992
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
Summer cooling driven by large volcanic eruptions over the Tibetan Plateau
1
2018
... 研究表明:赤道区域和北半球中低纬度的强火山喷发后1~2 a内,全球各地出现不同程度的降温,导致树轮密度偏低[16,97,100],因此常用树轮密度及其重建序列来研究火山喷发的降温效应.不同地区气候对火山喷发的响应存在差异,如高纬度地区对强火山喷发的响应要比为中纬度地区明显,延迟时间在1~2 a左右[101].北美西北部地区对火山喷发的响应较强,而北美的中部地区则响应偏弱[102].此外,在青藏高原地区树轮MXD中也发现了强的火山信号,青藏高原东南部在Tambora火山喷发之后有显著降温[79].青藏高原对中高纬度的火山爆发响应很弱,对热带火山爆发有显著的降温响应,但该降温响应有可能被厄尔尼诺的影响所抵消[103].总之,火山喷发对区域气候的影响与研究区地理位置、火山喷发的位置和强度有关[33]. ...
Underestimation of volcanic cooling in tree-ring-based reconstructions of hemispheric temperatures
2
2012
... 关于用树轮密度资料估计火山喷发带来的降温程度存在争议.Mann等[104]认为,火山喷发后的降温年份可能发生丢轮,从而低估了火山喷发带来的降温效应.但Esper等[105]假设欧洲北部和中部的MXD区域年表在1816年丢年,发现1722—1815年的MXD与夏季温度的相关系数分别从0.53、0.41降至0.02、0.03,从而否定了Mann等[104]的理论.也有研究认为光强减弱也能影响光合产物的合成,从而使密度偏低,故用密度重建的温度高估了火山喷发的降温效应[106].但Stoffel等[107]用树轮密度重建的温度变化与气候模式模拟的结果基本一致.Lea Schneider等[108]也指出树轮MXD受上个生长季树木生理条件的影响小,主要受当年生长季气候的影响,因此MXD能够较好地反映其区域在火山喷发后的降温程度.总之,丢轮、光照对树轮密度的影响,增加了重建温度的不确定性,降低了树轮密度对温度的代表性. ...
... [104]的理论.也有研究认为光强减弱也能影响光合产物的合成,从而使密度偏低,故用密度重建的温度高估了火山喷发的降温效应[106].但Stoffel等[107]用树轮密度重建的温度变化与气候模式模拟的结果基本一致.Lea Schneider等[108]也指出树轮MXD受上个生长季树木生理条件的影响小,主要受当年生长季气候的影响,因此MXD能够较好地反映其区域在火山喷发后的降温程度.总之,丢轮、光照对树轮密度的影响,增加了重建温度的不确定性,降低了树轮密度对温度的代表性. ...
Testing the hypojournal of post-volcanic missing rings in temperature sensitive dendrochronological data
1
2013
... 关于用树轮密度资料估计火山喷发带来的降温程度存在争议.Mann等[104]认为,火山喷发后的降温年份可能发生丢轮,从而低估了火山喷发带来的降温效应.但Esper等[105]假设欧洲北部和中部的MXD区域年表在1816年丢年,发现1722—1815年的MXD与夏季温度的相关系数分别从0.53、0.41降至0.02、0.03,从而否定了Mann等[104]的理论.也有研究认为光强减弱也能影响光合产物的合成,从而使密度偏低,故用密度重建的温度高估了火山喷发的降温效应[106].但Stoffel等[107]用树轮密度重建的温度变化与气候模式模拟的结果基本一致.Lea Schneider等[108]也指出树轮MXD受上个生长季树木生理条件的影响小,主要受当年生长季气候的影响,因此MXD能够较好地反映其区域在火山喷发后的降温程度.总之,丢轮、光照对树轮密度的影响,增加了重建温度的不确定性,降低了树轮密度对温度的代表性. ...
Temperature reconstructions from tree-ring densities overestimate volcanic cooling
2
2014
... 关于用树轮密度资料估计火山喷发带来的降温程度存在争议.Mann等[104]认为,火山喷发后的降温年份可能发生丢轮,从而低估了火山喷发带来的降温效应.但Esper等[105]假设欧洲北部和中部的MXD区域年表在1816年丢年,发现1722—1815年的MXD与夏季温度的相关系数分别从0.53、0.41降至0.02、0.03,从而否定了Mann等[104]的理论.也有研究认为光强减弱也能影响光合产物的合成,从而使密度偏低,故用密度重建的温度高估了火山喷发的降温效应[106].但Stoffel等[107]用树轮密度重建的温度变化与气候模式模拟的结果基本一致.Lea Schneider等[108]也指出树轮MXD受上个生长季树木生理条件的影响小,主要受当年生长季气候的影响,因此MXD能够较好地反映其区域在火山喷发后的降温程度.总之,丢轮、光照对树轮密度的影响,增加了重建温度的不确定性,降低了树轮密度对温度的代表性. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
Estimates of volcanic-induced cooling in the Northern Hemisphere over the past 1500 years
1
2015
... 关于用树轮密度资料估计火山喷发带来的降温程度存在争议.Mann等[104]认为,火山喷发后的降温年份可能发生丢轮,从而低估了火山喷发带来的降温效应.但Esper等[105]假设欧洲北部和中部的MXD区域年表在1816年丢年,发现1722—1815年的MXD与夏季温度的相关系数分别从0.53、0.41降至0.02、0.03,从而否定了Mann等[104]的理论.也有研究认为光强减弱也能影响光合产物的合成,从而使密度偏低,故用密度重建的温度高估了火山喷发的降温效应[106].但Stoffel等[107]用树轮密度重建的温度变化与气候模式模拟的结果基本一致.Lea Schneider等[108]也指出树轮MXD受上个生长季树木生理条件的影响小,主要受当年生长季气候的影响,因此MXD能够较好地反映其区域在火山喷发后的降温程度.总之,丢轮、光照对树轮密度的影响,增加了重建温度的不确定性,降低了树轮密度对温度的代表性. ...
Revising midlatitude summer temperatures back to A.D.?600 based on a wood density network
2
2015
... 关于用树轮密度资料估计火山喷发带来的降温程度存在争议.Mann等[104]认为,火山喷发后的降温年份可能发生丢轮,从而低估了火山喷发带来的降温效应.但Esper等[105]假设欧洲北部和中部的MXD区域年表在1816年丢年,发现1722—1815年的MXD与夏季温度的相关系数分别从0.53、0.41降至0.02、0.03,从而否定了Mann等[104]的理论.也有研究认为光强减弱也能影响光合产物的合成,从而使密度偏低,故用密度重建的温度高估了火山喷发的降温效应[106].但Stoffel等[107]用树轮密度重建的温度变化与气候模式模拟的结果基本一致.Lea Schneider等[108]也指出树轮MXD受上个生长季树木生理条件的影响小,主要受当年生长季气候的影响,因此MXD能够较好地反映其区域在火山喷发后的降温程度.总之,丢轮、光照对树轮密度的影响,增加了重建温度的不确定性,降低了树轮密度对温度的代表性. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
Tree growth and inferred temperature variability at the North American Arctic treeline
2
2009
... “分异性”问题是指基于树轮代用资料重建的温度序列在近几十年没有显著上升的趋势,与器测记录产生了显著差异[69,109].最早在阿拉斯加地区白云杉树轮宽度与密度对气候响应的研究中发现“分异性”问题[67].随后,在白令海峡与楚科奇地区[66]、芬诺斯坎迪亚北部[58]、比利牛斯山脉[63]等高纬度地区的树轮密度研究中也出现了“分异性”问题.中低纬度地区出现“分异性”问题的研究相对较少.在中国青藏高原东南部也存在该问题,Li等[79]发现在2005年后MXD与温度相关性降低. ...
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...
Reduced sensitivity of recent tree-growth to temperature at high northern latitudes
1
1998
... 关于造成“分异性”问题的原因,目前众说不一,主要的解释有3种.第一种观点认为“分异性”问题是由于低估降水影响造成的[27].近几十年来,高纬度地区的增暖大于低纬度地区,可能使高纬地区树轮生长的主要限制因子发生了改变,从而使温度与树木生长之间相关性降低[109].研究表明,近几十年来高纬地区树轮与降水的关系加强,干旱胁迫对树轮宽度与密度产生了很大的影响[63,67].第二种观点认为由于近些年来北半球高纬度地区植物可利用的光合有效辐射减少,树木径向生长缓慢,影响了树轮对升温的响应[106,110].第三种观点认为,标准化过程、空间分辨率、树种、树龄等因素影响了重建结果[62].不同树种、不同年龄阶段的树木生长趋势存在差异,而在建年表的过程中采用同一生长曲线拟合,再进行去趋势处理,可能导致重建温度与器测温度之间的偏差[73],如Grudd等[58]指出,当样本中加入了很多比较小树龄的样本时会影响生长趋势的拟合,最终影响重建结果;Schneider等[108]重建的北半球温度序列显示在近几十年没有显著的上升趋势,这可能是由于树轮密度空间分辨率过小引起的. ...



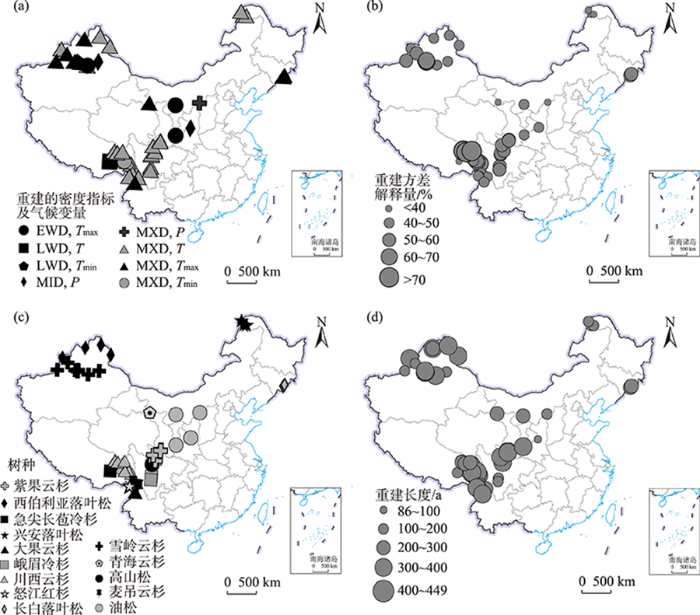


{kind=link}
{kind=link}
{kind=link}
{kind=link}