
Applications of chrysophyte stomatocysts in studies of aquatic environmental change
Received date: 2021-04-12
Request revised date: 2021-06-26
Online published: 2022-04-28
Supported by
National Natural Science Foundation of China(U20A2094)
National Natural Science Foundation of China(41572343)
Copyright
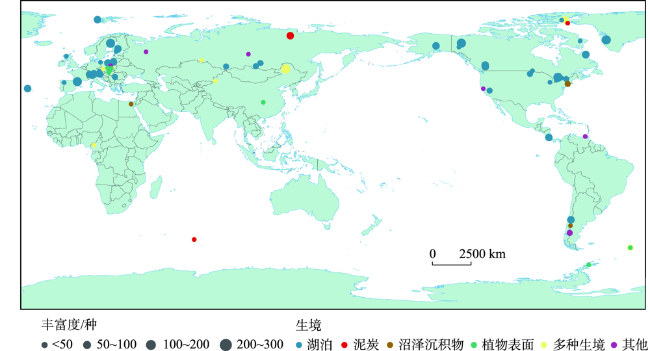
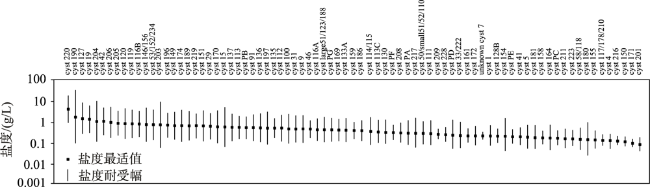
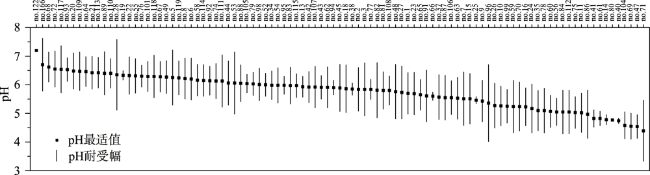
Stomatocysts are produced specifically by chrysophytes and distributed widely in freshwater environments, with siliceous shells well preserved in deposits. The main characteristics for the identification and classification of stomatocysts include pore, collar, annulus, and ornamentation. Because of their sensitivity to aquatic environmental changes, chrysophyte stomatocysts are broadly used in studies of aquatic environmental change, including reconstruction of paleotemperature and paleohydrology and assessment of lake eutrophication and acidification. Existing studies are mainly conducted in North America and Europe but few investigations exist in other regions. Compared with other bioindicators, the identification of stomatocysts is much more complex, probably limiting their broad application. Future studies should improve the identification and classification methods of stomatocysts in order to promote efficiency of identification and counting. Furthermore, modern investigation of stomatocysts should be strengthened to reveal global stomatocyst diversity and their ecological attributes, particularly in regions outside of Europe and North America. On these bases, applications of stomatocysts in the fields of peatland environmental change and biogeography should be strengthened.
BAI Xue , CHEN Xu . Applications of chrysophyte stomatocysts in studies of aquatic environmental change[J]. PROGRESS IN GEOGRAPHY, 2022 , 41(2) : 351 -360 . DOI: 10.18306/dlkxjz.2022.02.014
图1 鄂西泥炭地典型金藻孢囊扫描电镜(A~I)和光学显微镜照片(a~i)图中金藻泡囊名称如下:A、a:Stomatocyst 1, Duff & Smol;B、b:Stomatocyst 52, Duff & Smol;C、c:Stomatocyst 134, Duff & Smol;D、d:Stomatocyst 2, Bai & Chen;E、e:Stomatocyst 187, Brown & Smol;F、f:Stomatocyst 86, Duff & Smol;G、g:Stomatocyst 181, Brown & Smol;H、h:Stomatocyst 41, Pang & Wang;I、i:Stomatocyst 80, Hansen。 Fig.1 Micrographs of typical stomatocysts from peatlands of western Hubei Province under SEM (A-I) and LM (a-i) |
| [1] |
|
| [2] |
|
| [3] |
羊向东, 王荣, 董旭辉, 等. 中国湖泊古生态研究进展[J]. 湖泊科学, 2020, 32(5):1380-1395.
[
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
庞婉婷, 王全喜. 大兴安岭金藻孢囊图集 [M]. 北京: 科学出版社, 2017.
[
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
况琪军, 夏宜琤. 水体酸化对藻类影响的初步研究[J]. 环境科学学报, 1991, 11(4):387-392.
[
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}