姚永慧 , 张百平, 赵超
, 张百平, 赵超
YAO Yonghui, ZHANG Baiping, ZHAO Chao
通讯作者:
收稿日期: 2016-07-20
网络出版日期: 2017-04-20
版权声明: 2017 地理科学进展 《地理科学进展》杂志 版权所有
基金资助:
作者简介:
作者简介:姚永慧(1975-),女,湖北安陆人,博士,副研究员,研究方向为地理时空数据分析,E-mail:yaoyh@lreis.ac.cn。
展开
摘要
矮曲林是山地森林带在严酷的自然环境影响下发展演化而成的特殊植物群落,对中国山地植被和垂直带的研究具有重要的地理和生态意义。本文选择20世纪30年代以来在国内外文献中发表的52个矮曲林数据,结合Worldclim气候数据等,对中国矮曲林的类型和分布及其相应的气候条件进行了研究。结果表明:①中国的矮曲林主要包括山顶苔藓矮曲林、寒温性偃松矮曲林和岳桦矮曲林3类;其中,山顶苔藓矮曲林主要分布于秦巴山区以南的山地,而偃松矮曲林和岳桦矮曲林主要分布于东北地区。②山顶苔藓矮曲林主要由亚热带、热带地区的常绿阔叶林在山顶或山脊受山顶效应等的影响演化而来;而寒温性矮曲林主要由寒温性针叶林或针阔混交林在山顶或山脊条件下长期演化而来。山顶苔藓矮曲林自东向西随地势升高,其分布的海拔高度也逐渐升高;寒温性偃松矮曲林和岳桦矮曲林分布高度自南向北逐渐降低。③矮曲林分布在气候林线以下的山地,其气候条件足以支撑山地森林的发育,但由于山顶效应的存在(如强风、气候干冷或湿冷及土壤贫瘠等),只能发育矮曲林而非垂直地带性的山地森林。④中国东南部的很多山地高度小于3000 m,山顶效应明显,矮曲林广泛发育,造成了假林线的普遍存在。
关键词:
Abstract
Cripple trees that often lie between the upper montane forest and the alpine belt and normally grow at isolated summit locations or mountain ridges is a type of vegetation community shaped by the severe environmental conditions or the summit syndrome. The classification of cripple trees and their geographical distribution are important for the study of mountain altitudinal belts and mountain environment. Based on 52 cripple-tree datasets published in the literature since the 1930s and the climate data of Worldclim, this study analyzed the spatial distribution characteristics of cripple trees in China and their importance for identifying forest lines. The results show that: (1) Cripple trees in China are mainly consisted of mossy cripple trees and cold temperate cripple trees (including the Pinuspumila cripple trees and the Betulaermanii cripple trees). The Qinling-Daba Mountains are the boundary between mossy cripple trees and cold temperate cripple trees. Mossy cripple trees are distributed to the south of the Qinling-Daba mountain area, and cold-temperate cripple trees are distributed in northeast China. (2) With the increase of relief from east China to the west, the distribution altitude of mossy cripple trees increases gradually. The distribution altitude of mossy cripple trees on the southeast coast of China (Guangdong, Fujian, and Zhejiang) is about 800~1200 m, and west into the inland region such as west Zhejiang, Anhui, Jiangxi, Hubei, and Hunan, the altitude increases to about 1000~1800 m; in Chongqing, Guizhou, and Yunnan, the altitude increases to about 1500~3000 m, and higher than 3000~4500 m in the southeast of the Tibetan Plateau and the Himalayas. (3) The distribution altitude of the cold temperate cripple trees decreases from south to north. The distribution altitude of the Pinuspumila cripple trees decreases from 1400~1520 m at the Aokulidui Mountain to 1240 m at the Baikalu Mountain in the Greater Khingan Range; the altitude of the Betulaermanii cripple trees decreases from 1800~2100 m at the Changbai Mountain to 1100~1300 m at the Lesser Khingan Mountains. (4) The summit syndrome (strong winds, lower temperatures, and poor soils on the mountain summits or ridges) controls the formation of cripple trees. Mossy cripple trees are evolved from the subtropical montane evergreen broad-leaved forests or the tropical montane monsoon evergreen broad-leaved forests under the influence of the summit syndrome, and cold temperate cripple trees are evolved from the cold temperate coniferous forests or the coniferous and broad-leaved mixed forests in China under the influence of the summit syndrome. (5) The distribution altitude of cripple trees is often lower than the location of the forest climate line; their lower limits are not real forest line or climate forest line. Temperature in the warmest month (July) and annual precipitation for the distribution area of cripple trees (greater than 10°C and above 500 mm) verify that the climate condition of the cripple-tree distribution area can be suitable for forest growing. Most mountains in southeastern China are lower than 3000 m and cripple trees are present under the influence of the summit syndrome, therefore the forest lines in southeast China normally are not the real forest lines.
Keywords:
矮曲林(cripple tree或elfin forest/wood)一词最初出自德语“krummholz”(Schröeter, 1908; Hegi, 1958),随后被欧美植物学家广泛使用,用来描述落基山和欧洲山地林线上限附近矮小、变形的针叶树层或用来指森林和苔原的过渡带(Clausen, 1963; Wardle, 1965, 1973; Baig et al, 1976; Holtmeier, 1981)。矮曲林多分布在高山、亚高山的山顶或山脊,由于强烈而持续的大风、贫瘠的土壤、昼夜温差大、坡陡及冬春积雪较多,树木变形弯曲或矮化, 甚至匍匐地面、然后倾斜向上生长,形成耐瘠薄土壤、萌生力强,树种组成单一、呈单层外貌的矮曲林(吴征镒等, 1980)。矮曲林也常常被称为“山顶矮林”、“高山矮林”、“蟠蛇林”或“雾林”(叶居新, 1994),是在长期演替过程中发育成熟且相对稳定的植物群落类型。国外对矮曲林的研究相对较多,尤其是对阿尔卑斯山(Schröeter, 1908; Ellenberg, 1978, 1988)、落基山(Daubenmire, 1943; Marr, 1977)和科罗拉多山(Ives, 1973; Holtmeier, 1981)的矮曲林的物种组成、区系成分、生境特点和成因等进行了详细地研究。有些研究则详细对比了欧洲山地尤其是阿尔卑斯山与科罗拉多山矮曲林的区别,并对矮曲林的定义和名称进行了探讨(Holtmeier, 1981)。此外,在研究山顶效应(summit syndrome)或林线的分布时也多有提及矮曲林(Holtmeier, 1981; Körner et al, 2004; Holtmeier et al, 2005)。
国内对矮曲林专门系统的研究较少。根据《中国植被》(吴征镒, 1995)中矮曲林的物种组成、群落特征,中国矮曲林总体可分为寒温性矮曲林和亚热带山顶苔藓矮曲林2类。寒温性矮曲林主要分布在中国温带、寒温带地区如东北部地区的山顶或山脊, 以偃松矮曲林(Form. Pinus pumila)和岳桦矮曲林(Form. Betula ermanii)为代表。偃松矮曲林属寒温性针叶林植被型、寒温性常绿针叶林植被亚型、寒温性松林群系组。偃松矮曲林组成的树种极单纯, 仅在偃松丛间有呈团状分布的低矮的匍匐状的瘦桦(Betulo exilis)、兴安刺柏(Sabina Davurica)等,其林下与林间植物主要是各种地衣和藓类(吴征镒, 1995)。岳桦矮曲林属落叶阔叶林植被型、山地杨桦林植被亚型、桦木、桤木林群系组。岳桦矮曲林的组成极单纯,呈单层外貌,往往呈一株多干,倾斜生长, 树高平均为7~12 m;林内混生树种极少,仅伴生少量的花楸(Sorbus pohuashanensis)、东北赤杨(Alnus mandshurica)及鱼鳞云杉(Picea jezoensis)、长白山落叶松(Larix olgensis var. changpaiensis)和臭冷杉等;林下灌木稀少, 仅见散生的笃斯越桔(Vaccinum uliginosum)、西伯利亚刺柏(Juniperus sibirica)、蓝靓果(Loniceracaerulea var. edulis)、越桔(Vaccinum vitis-idaea)、牛皮杜鹃(Rhododendron xanthostephanum)等(吴征镒, 1995)。岳桦矮曲林因分布于高寒地带,很少有竞争者,且具有很强的有性和无性繁殖能力,可以占据云杉、冷杉所不能分布的生境,成为一种稳定的群落。
山顶苔藓矮曲林是中国亚热带山地常绿阔叶林和热带山地季风常绿阔叶林在山顶和山脊的环境条件下,自然演替的特殊群落变型,属常绿阔叶林植被型、山顶苔藓矮曲林植被亚型。其生境条件非常特殊,主要是山风强烈、日照少、气温低、气温的日变化大、云雾多、湿度大,土层浅薄,具有独特的群落学特征:①林木生长稠密、分枝低矮且粗壮;②叶型为小型叶或中型叶、革质且多茸毛;③小枝和叶片多具鳞片等旱生特征;④枝干或叶片上密被有附生的苔藓植物(吴征镒, 1995)。山顶苔藓矮曲林包括杜鹃矮曲林和吊钟花矮曲林2个群系组,其中杜鹃矮曲林包括猴头杜鹃、美丽马醉木、冷箭竹林(Form. Rhododendron simiarum, Pieris Formosa, Sinarundinaria fangiana), 猴头杜鹃、厚皮香林(Form. Rhododendron simiarum, Ternstroemia gymnanthera),大喇叭杜鹃、沙巴乌饭林(Form. Rhododendron excellens, Vaccinium chapaense),秀雅杜鹃矮曲林(Form. Rhododendron concinnum),都枝杜鹃矮曲林(Form. Rhododendron shanii)共5个群系;吊钟花矮曲林有吊钟花、锦杜鹃(Form. Enkianthus quinqueflorus, Thododendron fortunei)1个群系(吴征镒, 1995)。
在中国矮曲林的研究中,除对山顶苔藓矮曲林的物种组成、区系成分、基本生境特点、土壤种子库和生物量等有较详细的研究外(谢寿昌, 1987; 叶居新, 1994; 施济普, 2007; 徐捷等, 2010),对其他类型的矮曲林只是在植被分类或本底调查中略加描述(吴征镒等, 1980; 侯学煜, 1988; 杨小波等, 1994, 1995; 王映明, 1995a, 1995b; 彭华等, 1998),详细的研究很少。近年来,有些研究对中国山顶苔藓矮曲林的分布和特征进行了较全面的归纳和分析(徐捷等, 2010)。但对各类矮曲林在全国的分布和植物地理特征等方面的研究较少。由于矮曲林常分布在山地森林的上部边缘,是在具备发育山地森林的气温和降水条件下,山地森林带受山顶效应的影响而形成的一种特殊植被类型,对于山地植被垂直带、植被生态与环境研究等具有重要的生态意义,尤其对于解释中国东部林线分布特征具有重要的意义,因此有必要系统全面地研究中国矮曲林地理分布规律及其成因特征。
(1) 矮曲林数据。本文选择了52个自20世纪30 年代以来在国内外文献中发表的矮曲林数据,对中国矮曲林的分布特征进行研究。这些数据主要收集于以下书籍和论文:《Geoecology of the High Mountain Regions of Eurasia》(Troll, 1972),《珠穆朗玛峰地区科学考察报告》(郑度, 1975),《西藏植被》(中国科学院植物研究所, 1988),《云南植被》(吴征镒等, 1987),《中国植被地理及优势植物化学成分》(侯学煜, 1982),《中国植被》(吴征镒等, 1980),《广西大瑶山自然资源考察》(广西大瑶山自然资源综合考察队, 1988),“我国山地环境的自然特点及开发利用”(赵松乔, 1983),“湖北植被地理分布的规律性”(王映明, 1995a, 1995b),“略论川西山地的自然垂直带”(郑远昌等, 1984),“伏牛山森林群落物种多样性研究(一). 群落垂直分布与物种丰富度”(叶永忠等, 1999),“我国山地植被的垂直分布规律”(刘华训, 1981),“白石砬子山南北坡森林植被垂直分布的研究”(宋军等, 2005),“海南五指山森林的垂直分布及其特征”(符国瑷等, 1995),“广东乳阳八宝山自然保护区的植被特点”(张金泉, 1993)等文献和资料。矮曲林数据点覆盖青藏高原南部、云贵高原、秦岭及东部、东北地区(图1)。
(2) 气候数据本文所使用的气温和降水数据来自Worldclim数据集,下载自http://www.worldclim.org/。该数据集空间分辨率为1 km,为由Hijmans等(2005)根据全球气象台站观测数据(GHCN数据集、WMO CLINO数据集、FAOCLIM数据集及其他区域性、地方性的气候观测数据集),利用ANUSPLIN 软件中的“抿样条(the thin-plate smoothing splines)”内插方法内插的全球气温、降水数据集。文中矮曲林的气温和降水数据均取自该数据集。
首先,对于收集到的矮曲林数据,一部分有较精确的经纬度,或者根据山峰所在的位置能够获得精确的经纬度坐标;有些没有详细经纬度的数据,则按其分布区域,取区域经纬度的中间值。在上述52个矮曲林数据点中, 具有较精确经纬度坐标的数据共有21个,其中寒温性矮曲林8个,常绿苔藓矮曲林13个。然后,根据气温和降水数据、地形数据等,用ArcGIS软件及空间分析工具,计算和提取具有精确经纬度坐标的矮曲林数据点位置(21个数据点)的气温和降水信息、矮曲林带上下限数据等,在此基础上,对矮曲林的空间分布规律及其成因特征进行研究。
3.1.1 山顶苔藓矮曲林
根据所收集到的数据,山顶苔藓矮曲林主要分布于中国亚热带地区,包括青藏高原东南部和喜马拉雅山一带、云南、贵州、海南、广东、广西、湖南、湖北、浙江和安徽南部等南方大部分省区。本文山顶苔藓矮曲林的最北界为小秦岭主峰老鸦岔垴(图1),也有研究认为其最北界在伏牛山一带(徐捷等, 2010),因此其最北界应位于秦巴山区。由于山顶苔藓矮曲林是亚热带常绿阔叶林在山顶或山脊大风、多雾和低温等条件下长期演化形成的一种特殊植被类型, 其分布与中国亚热带常绿阔叶林和山地常绿阔叶林分布区相一致;并受地形分布的影响, 自东向西逐渐升高。在中国东南沿海(浙江、福建和广东),由于地势低平,山顶苔藓矮曲林的一般分布于海拔800~1200 m高度上;向西至中国第三级阶梯的内陆地区,如浙江西部、安徽、江西、湖北、湖南和广西,随着地势逐渐升高,山顶苔藓矮曲林一般于海拔1000~1800 m高度上;向西至第二阶梯的重庆、贵州和云南,山顶苔藓矮曲林的分布高度也继续升高,一般分布于海拔1500~3000 m;至第一阶梯藏东南地区及喜马拉雅山一带则基本分布在海拔3000~4500 m(图1)。
3.1.2 寒温性矮曲林
寒温性矮曲林主要分布于中国东北部的辽宁、吉林、黑龙江及内蒙古北部等寒温带地区。其中,偃松矮曲林是发育于寒温性针叶林(松林)上限附近的一种植被类型,在中国分布范围不大,主要分布于大兴安岭北部地区(吴征镒, 1995),且其分布高度自南向北逐渐降低,如在奥库里堆山分布于海拔1400~1520 m,向北至白卡鲁山则降低至海拔1240 m以上(图1);在长白山地区则常镶嵌于岳桦矮曲林之间。岳桦矮曲林分布于针阔混交林区域或落叶阔叶林上限附近,成为森林上限。在中国东北部分布广泛,尤其是在长白山上形成独特的岳桦林景观(吴征镒, 1995)。岳桦矮曲林自长白山向小兴安岭,其分布高度由南向北逐渐降低:在长白山一带基本上分布在海拔1800~2100 m(偃松矮曲林大概在海拔1900~2000 m),向北至张广才岭(大秃顶子、老秃顶子)一带则分布于海拔1180~1800 m,至小兴安岭白石砬子山则下降到海拔1100~1300 m(图1)。
3.2.1 矮曲林分布区的气候条件
从现有的寒温性矮曲林分布地点的气温和降水数据来看,寒温性矮曲林分布地点的最热月气温(7月)介于15~20℃,年降水量在500~800 mm之间, 基本上满足山地森林发育的气温和降水条件;但最冷月气温(1月)则非常低,通常在-20℃以下,年均温也基本上在0℃以下(-6.5~-0.2℃)(表1),生长季较短、冬季的气温较低。有研究表明,-10~-15℃的低温就会影响树干的发育(Wardle, 1971; Sakai, 1975),霜冻破坏极限(frost damage limit)和成熟极限(ripening limit)等都会影响林线的发育(Hutchins, 1947; Tuhkanen, 1980),再加上这些山顶的山风极其强烈、土壤条件瘠薄等因素的影响,树木为了适应这种恶劣的环境条件,逐渐发育成低矮的矮曲林。
表1 寒温性矮曲林最冷月、最热月及年平均气温、年降水量
Tab.1 Air temperature of the coldest month (January) and the warmest month (July), mean annual temperature, and annual precipitation for the cold temperate cripple trees
| 山地名称 | 经度/°E | 纬度/°N | 1月气温/℃ | 7月气温/℃ | 年均温/℃ | 年降水量/mm | 下限/m | 上限/m |
|---|---|---|---|---|---|---|---|---|
| 奥克里堆山 | 122.20 | 51.83 | -30.20 | 15.40 | -6.05 | 502 | 1400 | 1520 |
| 大兴安岭白卡鲁山 | 123.17 | 52.34 | -30.50 | 15.20 | -6.47 | 527 | 1240 | 1528 |
| 大兴安岭北端 | 123.90 | 51.95 | -28.70 | 16.80 | -4.68 | 505 | 1200 | 1300 |
| 长白山(东北北部) | 127.95 | 42.12 | -19.30 | 16.90 | -0.26 | 791 | 1800 | 2100 |
| 张广才岭大秃顶子 | 128.20 | 44.42 | -23.50 | 16.20 | -2.25 | 781 | 1400 | 1780 |
| 黄泥河自然保护区老白山南坡 | 128.24 | 44.05 | -22.80 | 16.00 | -2.09 | 780 | 1400 | 1650 |
| 老秃顶子自然保护区 | 128.36 | 44.38 | -21.50 | 18.20 | -0.20 | 715 | 1180 | 1250 |
| 小兴安岭白石砬子 | 128.96 | 46.91 | -23.40 | 19.60 | -0.08 | 670 | 1100 | 1270 |
本文收集的山顶苔藓矮曲林数据中,具有较精确经纬度坐标的共13个,主要分布于中东部地区(第三级阶梯的内陆地区),遗憾的是云贵高原及青藏高原的矮曲林数据基本上没有记录精确的地理位置。根据这13个山顶苔藓矮曲林分布地点的气温来看(表2),最热月气温(7月)介于17~22℃之间;最冷月气温(1月)除海南、广东、广西和江浙等沿海一带能达到4~12℃外,其他地方的1月气温基本上都低于0℃;山顶苔藓矮曲林分布区的年平均气温要低于所在省份的年均气温,除海南、广东、广西和江浙一带基本上在10℃以上(10~17℃)外,其他地区均低于10℃(5~10℃)。从年降水量上来看,除秦岭南部低于1000 mm(600~1000 mm),其他地区的年降水量基本上在1500~2300 mm之间。已有的研究表明,亚热带常绿阔叶林的上限常常与最冷月均温-2~1℃等温线相当(Wolfe, 1980; Ohsawa, 1990),最冷月气温对常绿阔叶林上限分布具有重要的影响,这也是这些山顶发育了苔藓矮曲林的一个重要原因。同寒温性矮曲林的气温和降水量相比,山顶苔藓矮曲林分布区的气温略高,但降水量较大。湿冷多雾、强风、土壤贫瘠是山顶苔藓矮曲林形成的重要影响因素(吴征镒, 1995)。
表2 山顶苔藓矮曲林最冷月、最热月及年平均气温、年降水
Tab.2 Air temperature of the coldest month (January) and the warmest month (July), mean annual temperature, and annual precipitation for the mossy cripple trees
| 山地名称 | 经度/°E | 纬度/°N | 1月气温/℃ | 7月气温/℃ | 年均温/℃ | 年降水量/mm | 下限/m | 上限/m |
|---|---|---|---|---|---|---|---|---|
| 海南五指山 | 109.70 | 18.90 | 12.3 | 20.4 | 17.13 | 2017 | 1400 | 1867 |
| 广西大瑶山圣堂山 | 110.11 | 23.96 | 5.8 | 22.1 | 14.79 | 1628 | 1500 | 1979 |
| 广东乳阳八宝山 | 113.13 | 24.43 | 6.8 | 24.0 | 16.01 | 1761 | 1600 | 1700 |
| 武夷山主峰黄岗山 | 117.78 | 27.86 | 0.1 | 19.0 | 9.57 | 2293 | 1750 | 2000 |
| 贵州梵净山 | 108.69 | 27.92 | -2.6 | 17.2 | 7.52 | 1525 | 2240 | 2400 |
| 浙江北山 | 119.91 | 29.06 | 4.0 | 27.3 | 15.70 | 1549 | 800 | 1310 |
| 湖北星斗山 | 109.13 | 30.03 | 1.2 | 22.0 | 11.80 | 1524 | 1600 | 1751 |
| 安徽清凉峰 | 118.86 | 30.10 | -2.4 | 19.4 | 8.72 | 1969 | 1580 | 1730 |
| 安徽黄山光明顶 | 118.16 | 30.13 | -2.4 | 18.6 | 8.46 | 2229 | 1500 | 1650 |
| 浙江龙王山 | 119.40 | 30.38 | -1.7 | 21.1 | 9.83 | 1681 | 1400 | 1587 |
| 河南伏牛山玉皇顶 | 111.82 | 33.64 | -5.5 | 18.2 | 6.68 | 974 | 1900 | 2100 |
| 河南伏牛山老君山 | 111.64 | 33.72 | -6.5 | 17.1 | 5.54 | 999 | 2000 | 2200 |
| 小秦岭老鸦岔脑 | 111.40 | 34.40 | -1.8 | 24.9 | 12.14 | 604 | 1900 | 2000 |
3.2.2 山顶效应是矮曲林发育的重要影响因素
无论是寒温性矮曲林,还是亚热带山顶苔藓矮曲林,绝大部分矮曲林分布在本来可以发育山地森林带的地方,但由于山顶或山脊的大风与贫瘠陡峭的地表,因而只能发育矮曲林。从表1和表2中最热月(7月份)气温和年降水量来看,无论是寒温性矮曲林还是山顶苔藓矮曲林发育的山顶,最热月平均气温都远远大于10°C,年降水量>500 mm,可满足森林发育的热量和水分条件(Troll, 1973),但都没有进一步发育山地森林。除最冷月气温和年均温较低的限制和影响外,山顶效应在矮曲林的形成和发育中起着重要的作用(吴征镒等, 1980; Leuschner, 1996; Holtmeier et al, 2005)。首先,这些相对孤立的山顶或山脊,山风极其强烈,树木多匍匐在地生长或形成旗树;其次,这些山顶的土壤非常贫瘠,不仅土层薄,而且土壤中的有机质含量和矿物质等也非常少(Grubb, 1971);此外,中国亚热带山地多雾和日照少、湿度大,也是山顶苔藓矮曲林发育的重要影响因素。因此,矮曲林在生态学上标志着山地森林带的上限,但其下限并不是真正的林线或气候林线,而是指由于山体的高度或山顶效应的影响,出现的一种特殊的植物群落,也可称为假林线,具有重要的生态意义。
从植物区系和演化来看,山顶苔藓矮曲林是中国亚热带山地常绿阔叶林和热带山地季风常绿阔叶林在山顶效应这种极端环境下长期演化而来;而寒温性矮曲林则是中国寒温性针叶林或针阔混交林等在山顶效应的环境性逐渐演化形成的植物群落。总的来讲,矮曲林是地带性植被在山顶效应这种非地带性因素的影响下形成的一种特殊植被类型,也有人称其为地形顶极、半地带性的植被类型(叶居新, 1994)。矮曲林的出现也表征了山顶效应的存在,在地理学上具有重要的研究意义。另外,由于矮曲林分布的海拔较高,较少受到的人类干扰,多数群落保留了较原始的状态,这为研究中国山地植被的基本分布和生态等提供了一个很好的对象。
3.2.3 矮曲林对中国东南部山地林线研究的意义
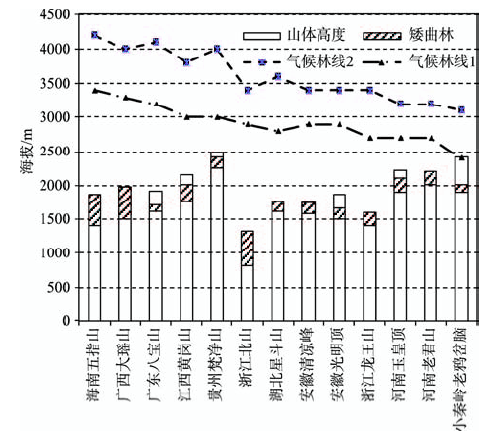
根据已有的研究和文献资料,中国东南部山地的林线除台湾玉山的林线高度可达3600 m外,一般都在2000 m以下,个别山地的林线能达到2000 m以上。但按照气温和降水条件,中国东南部的气候林线可以达到2200~3600 m(张百平等, 2015),部分研究估算的中国东南部的气候林线可达3000~4000 m(蒋复初等, 2004),但实际林线远低于气候林线(图2)。中国东南部地区由于处于东南季风影响区,并且很多山地高度小于3000 m,山顶效应作用明显,矮曲林发育, 造成了假林线的普遍存在。这也可从另一个侧面解释了为何中国东南部林线远远低于西南部的林线高度。
图2 中国东南部部分山地的气候林线及矮曲林分布高度(气候林线1引自张百平等(2015),气候林线2引自蒋复初等(2004))
Fig.2 Climate forest line and the distribution altitude of cripple-tree forest for mountains in southeastern China(climate forest line 1 was cited from
(1) 中国的矮曲林主要包括山顶苔藓矮曲林和寒温性偃松矮曲林和岳桦矮曲林;其中,秦巴山区是一条重要的分界线,以南的广大亚热带、热带山地主要分布山顶苔藓矮曲林,而偃松矮曲林和岳桦矮曲林则主要分布于东北地区。秦巴山区是中国亚热带和温带的分界线,再向北,由于气温较低, 因而限制了山顶苔藓矮曲林向北发展。
(2) 山顶苔藓矮曲林主要由亚热带、热带地区的常绿阔叶林在山顶或山脊受山顶效应等的影响演化而来;而寒温性矮曲林(偃松矮曲林和岳桦矮曲林)则主要由寒温性针叶林或针阔混交林在山顶或山脊条件下长期演化而来。山顶苔藓矮曲林自东向西随地势升高,其分布的海拔高度也逐渐升高, 寒温性偃松矮曲林和岳桦矮曲林分布的海拔高度自南向北逐渐降低。
(3) 山顶效应即山顶或山脊强风、较低的气温及贫瘠的土壤等生境条件,是矮曲林形成的主导因素。无论是热带、亚热带山顶苔藓矮曲林,还是寒温性偃松矮曲林和岳桦矮曲林,都是在山脊或山顶地带,由于山风大且气温低、土层瘠薄、岩石裸露的生境条件下,由适应山顶这种严酷生存条件的树种经过长期自然演化而形成的。
(4) 矮曲林的分布位置远低于气候林线,其下限并非真正的林线或气候林线。矮曲林分布区的年降水和最热月气温都表明其环境条件是可以发育森林的(年降水量>500 mm,最热月平均气温都远远大于10°C)。中国东南部的很多山地高度小于3000 m,山顶效应作用明显,矮曲林发育,造成了假林线的普遍存在。
(1) 矮曲林形成的生境非常特殊,山顶效应起着非常重要的作用,但由于局地的气候因素难以获取,相关研究较少。目前,随着观测手段和仪器设备的提升,有条件进行较为详细的调查研究。首先,通过测风仪、测温仪、土壤水分自动监测系统等,可以获取包括风速、气温、日照、湿度、土壤条件等的观测数据;另外,由于遥感热红外技术的发展,为快速精确地获取地表温度、降水等创造了条件,尤其是将调查观测数据与遥感热红外数据相结合,可以反演矮曲林发育的气候环境条件,从而对矮曲林发育环境进行详细研究。同时,山顶效应的影响研究,对于揭示中国东部林线的分布特征具有非常重要的意义,也是自然地理学非地带性作用的重要研究内容。
(2) 本文所用的数据均来源于已发表的文献资料,由于一部分数据并没有提供精确的经纬度坐标,可能在空间定位上有些误差,但从宏观区域尺度上来看,所得到结果仍可反映矮曲林在中国的总体分布趋势。此外,由于收集到的数据多来自垂直带谱研究的相关文献资料,本文没有详细地研究矮曲林的物种组成、植物的区系组成等群落特征。另外,文献数据基本上都是20世纪80年代以前的调查数据,在人类活动和气候变化的影响下,矮曲林的分布也相应地发生变化,迫切需要最新的详细调查数据加以补充。尤其是在数据比较稀缺的西部地区以及2类矮曲林的过渡带地区(秦巴山区)更应该开展详细调查,获取最新的矮曲林分布数据。鉴于矮曲林生境的独特性和分布的特殊性,在详细调查的基础上,只有对其生境和分布规律进行系统、深入地研究,才能进一步揭示矮曲林的生态意义。
The authors have declared that no competing interests exist.
| [33] |
Alpenflora [M]. |
| [34] |
Very high resolution interpolated climate surfaces for global land areas [J]. |
| [35] |
What does the term 'krummholz' really mean? Observations with special reference to the Alps and the Colorado Front Range [J].https://doi.org/10.2307/3673062 URL [本文引用: 1] 摘要
ABSTRACT The term "Krummholz" is applied to the bush-like growth-forms of Pinus mugo prostrata, Alnus viridis, and some other species which are genetically determined. The term has also long been used by English-speaking botanists to refer to the climatically stunted and distorted trees in the forest-tundra ecotone. Thus, distinct ecological and plant-geographical differences are disguised which, in a more precise differentiation of the mountain plant cover, ought to be accentuated. The author tries to explain this by observation and field studies in the European high mountains and in the Rocky Mountains and suggests restricting the German term "Krummholz" to the genetically controlled shrub-like species; the timberline species shaped by climatic influences should be called cripple-trees. Thus, the elfin wood is nothing but a cripple facies of the upper montane forest. /// En allemand, on appelle "Krummholz" les espèces de bois comme Pinus mugo prostrata et Alnus viridis qui sont d'une pousse tordue par voie d'hérédité. En anglais, on appelle "Krummholz" depuis quelque temps aussi les formes des arbres à la limite forestière supérieure qui sont occasionées par le climat. Avec ce transfert, des différences importantes en vue de l'écologie et de la géographie floristique sont effacées qui devront être soulignées pour obtenir la différenciation la plus exacte de la végétation de haute montagne. Cela, l'auteur essaye de démontrer au moyen des observations comparatives dans les hautes montagnes européennes et dans les montagnes rochenses. Il suggère de réserver le terme "Krummholz" pour les espèces de pousse tordue fixée genetiquement et d'appeler arbres deformés les formes de pousse tordue forcées par les constraints du climat. Alors, le "elfin wood" n'est qu'une forme tordue de la forêt près de sa limite supérieure. /// Als Krummholz werden im Deutschen die erblich krummwüchsigen Holzarten wie Pinus mugo prostrata und Alnus viridis bezeichnet. Im Englischen ist es seit geraumer Zeit üblich geworden, auch die klimatisch bedingten Wuchsformen im Waldgrenzbereich "Krummholz" zu nennen. Mit dieser 05bertragung werden aber wesentliche 02kologische und pflanzengeographische Unterschiede verwischt, die im Hinblick auf eine m02glichst genaue Differenzierung der Hochgebirgsvegetation eigentlich hervorgehoben werden mü08ten. Dies versucht der Autor anhand vergleichender Beobachtungen und Studien in europ01ischen Hochgebirgen und in den Rocky Mountains zu begründen. Er schl01gt vor, den Terminus "Krummholz" den Arten mit genetisch fixierter Krummwüchsigkeit vorzubehalten und die klimatisch induzierten Zwangswuchsformen als Krüppelwuchsformen oder Baumkrüppel zu bezeichnen.
|
| [36] |
Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales [J].https://doi.org/10.1111/j.1466-822X.2005.00168.x URL 摘要
ABSTRACT The sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change are increasingly discussed in terms of climate change, often forgetting that climate is only one aspect of environmental variation. As treeline heterogeneity increases from global to regional and smaller scales, assessment of treeline sensitivity at the landscape and local scales requires a more complex approach than at the global scale. The time scale (short-, medium-, long-term) also plays an important role when considering treeline sensitivity. The sensitivity of the treeline to a changing environment varies among different types of treeline. Treelines controlled mainly by orographic influences are not very susceptible to the effects of warming climates. Greatest sensitivity can be expected in anthropogenic treelines after the cessation of human activity. However, tree invasion into former forested areas above the anthropogenic forest limit is controlled by site conditions, and in particular, by microclimates and soils. Apart from changes in tree physiognomy, the spontaneous advance of young growth of forest-forming tree species into present treeless areas within the treeline ecotone and beyond the tree limit is considered to be the best indicator of treeline sensitivity to environmental change. The sensitivity of climatic treelines to climate warming varies both in the local and regional topo-graphical conditions. Furthermore, treeline history and its after-effects also play an important role. The sensitivity of treelines to changes in given factors (e.g. winter snow pack, soil moisture, temperature, evaporation, etc.) may vary among areas with differing climatic characteristics. In general, forest will not advance in a closed front but will follow sites that became more favourable to tree establishment under the changed climatic conditions.
|
| [37] |
The bases for temperature zonation in geographical distribution [J].https://doi.org/10.2307/1948663 URL [本文引用: 3] 摘要
See full-text article at JSTOR
|
| [38] |
Studies in high altitude Geoecology of the Colorado Front Range: A review of the research program of the Institute of Arctic and Alpine Research, University of Colorado [J].URL 摘要
The past, present, and future research programs of the Institute of Arctic and Alpine Research (INSTAAR) are described and discussed. The early phase (1950 to 1967) concentrated upon reconnaissance, establishment and maintenance of climatological stations, and tabulation of data as a basis for the study of the geoecology of the high mountains of Colorado with emphasis on the Front Range where INSTAAR's mountain research station is located at 2,930 m elevation. The impact of significant expansion in 1968 in the size, scope, and extent of INSTAAR activities to the present is discussed. The climatological data is being analyzed using the TAXIR System of data reduction. Biological studies, many of which are associated with the IBP Tundra Biome's alpine site on Niwot Ridge, include tundra plant productivity, tree line fluctuations in the forest-tundra ecotone, ornithology, and entomology, and others. The abiotic field studies include glacier mass balance, soil movement, geochronology using lichenometry, stratigraphy, palynology, and mapping of tills and moraines. Lately application of remote sensing using NASA/ERTS-1 and Skylab imagery has widened the possible scope of landscape studies. Future research will develop work to date and apply it to resource development problems and conservation. In the short-term a monograph on the geoecology of the Colorado Front Range is planned. The paper concludes with a list of cited references pertaining to Front Range research and a bibliography of related publications and current master's degree and doctoral dissertation research topics.
|
| [39] |
A world-wide study of high altitude treeline temperatures [J].https://doi.org/10.1111/j.1365-2699.2003.01043.x URL [本文引用: 1] 摘要
ABSTRACT Aim At a coarse scale, the treelines of the world's mountains seem to follow a common isotherm, but the evidence for this has been indirect so far. Here we aim at underpinning this with facts.Location We present the results of a data-logging campaign at 46 treeline sites between 68° N and 42° S.Methods We measured root-zone temperatures with an hourly resolution over 1–3 years per site between 1996 and 2003.Results Disregarding taxon-, landuse- or fire-driven tree limits, high altitude climatic treelines are associated with a seasonal mean ground temperature of 6.7 °C (±0.8 SD; 2.2 K amplitude of means for different climatic zones), a surprisingly narrow range. Temperatures are higher (7–8 °C) in the temperate and Mediterranean zone treelines, and are lower in equatorial treelines (5–6 °C) and in the subarctic and boreal zone (6–7 °C). While air temperatures are higher than soil temperatures in warm periods, and are lower than soil temperatures in cold periods, daily means of air and soil temperature are almost the same at 6–7 °C, a physics driven coincidence with the global mean temperature at treeline. The length of the growing season, thermal extremes or thermal sums have no predictive value for treeline altitude on a global scale. Some Mediterranean (Fagus spp.) and temperate South Hemisphere treelines (Nothofagus spp.) and the native treeline in Hawaii (Metrosideros) are located at substantially higher isotherms and represent genus-specific boundaries rather than boundaries of the life-form tree. In seasonal climates, ground temperatures in winter (absolute minima) reflect local snow pack and seem uncritical.Main conclusions The data support the hypothesis of a common thermal threshold for forest growth at high elevation, but also reflect a moderate region and substantial taxonomic influence.
|
| [40] |
Timberline and alpine vegetation on the tropical and warm-temperate oceanic islands of the world: Elevation, structure and floristics [J].https://doi.org/10.1007/BF00118271 URL 摘要
Abstract In the oceans of the tropical and warm-temperate zone (40掳 N40掳 S), only a small number of islands are high enough to show timberline and alpine vegetation. Excluding large islands with a more continental climate, only the following oceanic islands are relevant: Pico (Azores), Madeira, Tenerife, Gran Canaria and La Palma (Canary islands), Fogo (Cape Verde islands), Fernando Poo (Bioko) and Tristan da Cunha in the Atlantic Ocean, R茅union and Grande Comore (Ngazidja) in the Indian Ocean, Yakushima (Japan), Maui and Hawaii (Hawaiian islands), and Mas Afuera (Juan Fernandez islands) in the Pacific Ocean. Timberline and alpine vegetation exist here under a unique combination of a highly oceanic climate and a marked geographic isolation which contrasts with the tropical alpine vegetation in the extended mountains of South America, Africa and Southeast Asia. This review seeks to identify common physiognomic patterns in the high elevation vegetation that exist despite the fact that the islands belong to different floristic regions of the world. Based on the existing literature as well as personal observation, an overview of the elevation, physiognomy and floristics of the forest (and tree) line and the alpine vegetation on 15 island peaks is given. The forest line ecosystems are dominated either by conifers (Canary islands, Yakushima), heath woodland (Azores, Madeira, R茅union, Grande Comore, Fernando Poo) or broad-leaved trees (Hawaiian islands, Juan Fernandez islands, Tristan da Cunha). In the subalpine and alpine belts, dry sclerophyllous scrub occurs on island mountains that are exposed to the trade winds (Canary islands, Cape Verde islands, Hawaiian islands, R茅union, Grande Comore). These peaks are more or less arid above the forest line because a temperature inversion restricts the rise of humid air masses further upslope. In the summit regions of the remaining islands, which are located either in the wet equatorial and monsoonal regions or in the temperate westerly zones without an effective inversion layer, mesic to wet vegetation types (such as grassland, alpine heathland and fern scrub) are found. Compared to mountains at a similar latitude in continental areas, the forest line on the islands is found at 1000 to 2000 m lower elevations. The paper discusses four factors that are thought to contribute to this forest line depression: (1) drought on trade-wind exposed island peaks with stable temperature inversions, (2) the absense of well-adapted high-altitude tree species on isolated islands, (3) immaturity of volcanic soils, and (4) an only small mountain mass effect that influences the vertical temperature gradient.
|
| [41] |
The development and movement of tree islands near the upper limit of yree growth in the Southern Rocky mountains [J].https://doi.org/10.2307/1936937 URL [本文引用: 1] 摘要
Dwarfed and deformed plants of tree species occur as 'tree islands' in a matrix of tundra vegetation near the upper limit of tree growth in the Southern Rocky Mountains. Many of these islands are asymmetrical, having a surface layer of dead branches to windward and some live branches to leeward. I made repeated observations of these islands in all seasons from 1956 to 1967 along a 1.75-km belt ?20 m wide in Niwot ridge in the Front Range was of Denver, Colorado. The location of islands was mapped on a transparent overlay on an aerial photograph, annual growth of stem tips was measured, and an abundance of notes and photographs was taken. Some tree islands of Engelmann spruce (Picea engelmannii) and subalpine fir (Abies lasiocarpa) are conspicuous because they grow in microsites in which no tree seedlings have been found, and which general observations suggest would be outside their environmental amplitude. These islands often have live branches connected to windward with dead branches, some of which have roots. These observations led to the hypothesis that some tree islands move along the ground climbing out of more moderate microsites in which a seed germinated and moving into microsites for which they are not well adapted. In this movement process, wind-exposed organs are killed, while leeward stems grow to leeward often taking root and forming an upright stem system in the process of 'stem tip layering.' Islands with interconnected live and dead stems to windward, illustrating movements of 5 m, are common; isolated dead stumps and wood fragments in a line extending to windward indicate that some islands may have moved at least 15 m. The origin of tree islands, support for the movement hypothesis, and other features of tree island ecology and discussed.
|
| [42] |
An interpretation of latitudinal patterns of forest limits in South and East Asian mountains [J].https://doi.org/10.2307/2261115 URL [本文引用: 1] 摘要
A discussion of forest limits (treelines) in S. and E. Asia in relation to floristics (mainly evergreen conifers with some deciduous broadleaves in northern temperate areas, but evergreen broadleaves on tropical mountains), temperature conditions (seasonality, minimum temperature affecting winter survival, minimum temperature sums affecting summer growth and reproduction), and altitude and lati...
|
| [43] |
Freezing resistance of evergreen and deciduous broad-leaf trees in Japan with special reference to their distributions [J].
Dormant one-year-old twigs collected from mature trees growing under different climates during mid-winter were artificially hardened at sub-freezing temperatures to overcome the differences of the sites of collection and to induce maximum freezing resistance. Evergreen broad leaf trees which range from the tropics in East Asia to the warm Pacific sea coasts can survive freezing to-5--7℃. Hardy evergreen broad-leaf trees and less hardy deciduous trees which have their northern limits of natural ranges in the northern Kanto District resistered freezing from -10 to -15℃. In these trees, the winter minimal temperatures appear to be the principal factor limiting the northern limits of their natural ranges. Most of the hardy deciduous trees which have their northern limits of natural ranges in northern Honshu and southern Hokkaido resisted freezing to only about -30℃. In these genera, the winter minimal temperatures appear to be the principal factor governing their geographical distribution in severe cold climates. In most of the more widely distributed deciduous species which extend to inland Hokkaido, Sakhalin and East Manchuria, the buds and cortical tissues of the twigs resisted freezing to -70℃.
|
| [44] |
Das pflanzenleben der Alpen: Eine schilderung der hochgebrigsflora [M].
|
| [1] |
海南五指山森林的垂直分布及其特征 [J].
本文探讨了海南五指山森林垂直分布其特征,并根据五指山自然保护区的6000m^2样地调查 材料,连同区内一些路线调查材料统计结果,计有维管束植物998种,576属,178科,按照植物地理成份分析,指出以樟科、桃金科、壳斗科、棕榈科等 科、属、种为主的五指山热带、亚热带植物属占总属的89%是很高的。
The vertical distribution and characteristics of forest of Wuzhishan, Hainan [J].
本文探讨了海南五指山森林垂直分布其特征,并根据五指山自然保护区的6000m^2样地调查 材料,连同区内一些路线调查材料统计结果,计有维管束植物998种,576属,178科,按照植物地理成份分析,指出以樟科、桃金科、壳斗科、棕榈科等 科、属、种为主的五指山热带、亚热带植物属占总属的89%是很高的。
|
| [45] |
The three-dimensional zonation of the Himalayan system [M] |
| [46] |
The upper timberlines in different climatic zones [J].URL 摘要
In the past the upper timberlines and their ecological causality were mostly studied in high mountains of the humid temperate zones of the Northern Hemisphere with their strong thermal contrasts of summer and winter. They are generally determined by the duration of certain summer temperature values and in their topoclimatic differentiation controlled by the accumulation and deflation of snow. But they are not climatically equivalent, not even in a relatively small mountain system such as the Alps or the Tatra mountains from what is shown by the change in the limit-forming trees (spruce, larch, pine, fir, beech, birch, etc.). The upper timberlines in the humid tropics are completely different in physiognomy, life forms, climatic conditions, and topoclimatic effects; they are without seasonal variations of temperatures. In most cases they are formed by a dense evergreen forest with dozens of broad-leaved trees, sometimes by a fringing woodland (Polylepis, Ericacea, Hagenia, Leptospermum). In the arid belts, which extend from tropical to cold temperate latitudes, where the forest belts have a lower and an upper limit ("girdle forests"), also the upper timberline can be caused, at least in part, by aridity factors. In the Mediterranean area and in the Middle East we distinguish a sub-Mediterranean subzone with upper timber belts of boreal types, a fully Mediterranean subzone with specific Mediterranean trees as uppermost timber belt (Quercus ilex, Q. tozza, Pinus leucodermis, Cedrus atlantica), and a southerly Mediterranean steppe belt where generally juniper species are upper limiting trees. The timberlines in the cool temperature zone of the Southern Hemisphere with its high oceanity show more affinity to the tropical highlands than to the boreal zones.
|
| [2] |
|
| [47] |
Climatic parameters and indices in plant geography [M]. |
| [48] |
A comparison of alpine timber lines in New Zealand and North America [J].https://doi.org/10.1080/0028825X.1965.10876989 URL 摘要
. New Zealand Journal of Botany 4:1, pages 42-56.
|
| [3] |
|
| [49] |
An explanation for alpine timberline [J]. |
| [50] |
New Zealand timberlines [J].
|
| [4] |
|
| [5] |
中国大陆森林线空间分布特征及其与多年冻土线、气候雪线的关系 [J].https://doi.org/10.3969/j.issn.1006-6616.2004.04.001 URL [本文引用: 1] 摘要
在自然植被垂直带谱上一般以暗针叶林带的上限代表森林线,它的海拔高度取决于温度及其年较差。本文主要根据我国一些地区现代暗针叶林带上限的海拔高程和102个气象台站的气象资料,计算出我国的森林线的理论海拔高度,并编绘出我国的森林线高程等值线图以及多年冻土线与森林线高差等值线图和现代气候雪线与森林线高差等值线图。在此基础上研究了我国森林线的空间分布特征及其与多年冻土线、气候雪线的关系。
Features of space distribution of the forest line and relations between the forest line and climatic limit of permafrost and climatic snowline in China [J].https://doi.org/10.3969/j.issn.1006-6616.2004.04.001 URL [本文引用: 1] 摘要
在自然植被垂直带谱上一般以暗针叶林带的上限代表森林线,它的海拔高度取决于温度及其年较差。本文主要根据我国一些地区现代暗针叶林带上限的海拔高程和102个气象台站的气象资料,计算出我国的森林线的理论海拔高度,并编绘出我国的森林线高程等值线图以及多年冻土线与森林线高差等值线图和现代气候雪线与森林线高差等值线图。在此基础上研究了我国森林线的空间分布特征及其与多年冻土线、气候雪线的关系。
|
| [6] |
我国山地植被的垂直分布规律 [J].https://doi.org/10.11821/xb198103003 URL 摘要
正地球上植被的大型分布,明显地呈现地带性变化。平地上表现为水平地带性,包括纬度地带性和干燥度地带性;山地上表现为垂直(地)带性,就是所谓“三维空间”分布现象。我国地广山多,自然条件复杂,研究山地植被的分布规律不但具有理论意义,而且也富有实践意义。
The vertical zonation of mountain vegetation in China [J].https://doi.org/10.11821/xb198103003 URL 摘要
正地球上植被的大型分布,明显地呈现地带性变化。平地上表现为水平地带性,包括纬度地带性和干燥度地带性;山地上表现为垂直(地)带性,就是所谓“三维空间”分布现象。我国地广山多,自然条件复杂,研究山地植被的分布规律不但具有理论意义,而且也富有实践意义。
|
| [7] |
无量山中山湿性常绿阔叶林及其植物区系的初步研究 [J].
无量山中山湿性常绿阔叶林是其垂直带上最具特征性的植被类型。从这类森林中231种优势和常见植物的种类结构分析看,只有热带亚洲成分、东亚中的中国-喜马拉雅成分、和中国特有成分贯穿乔木A、B层,灌木层,草本层和层外植物几大类中;而中国特有种的进一步分析表明了它的区系成分的亚热带性质,温带性质不是很显著。该地这类森林与云南同类型植被有诸多方面的相似性,但是大致上每一个大的山头有一套植物种类成分,而以各种石栎树种加以区分,其它重要伴生种及伴生种的各类组合千差万别。对这类森林中35个特有种的分析显示区系平衡点邻近范围有较大的物种分化水平;它们所隶属的属的分析表明,这类植被的温带性质又较该地整体水平为强;它们的近缘种分布区域对无量山此类森林与这些区域相应类型植被的区系联系和分化有一定指示意义
The preliminary floristical study on mid-montane humid evergreen broad-leaved forest in Mt. Wuliangshan [J].
无量山中山湿性常绿阔叶林是其垂直带上最具特征性的植被类型。从这类森林中231种优势和常见植物的种类结构分析看,只有热带亚洲成分、东亚中的中国-喜马拉雅成分、和中国特有成分贯穿乔木A、B层,灌木层,草本层和层外植物几大类中;而中国特有种的进一步分析表明了它的区系成分的亚热带性质,温带性质不是很显著。该地这类森林与云南同类型植被有诸多方面的相似性,但是大致上每一个大的山头有一套植物种类成分,而以各种石栎树种加以区分,其它重要伴生种及伴生种的各类组合千差万别。对这类森林中35个特有种的分析显示区系平衡点邻近范围有较大的物种分化水平;它们所隶属的属的分析表明,这类植被的温带性质又较该地整体水平为强;它们的近缘种分布区域对无量山此类森林与这些区域相应类型植被的区系联系和分化有一定指示意义
|
| [8] |
云南山顶苔藓矮林群落生态学与生物地理学研究[D] .
Community ecology and biogeography of the mossy dwarf forest in Yunnan[D].
|
| [9] |
白石砬子山南北坡森林植被垂直分布的研究 [J].https://doi.org/10.3969/j.issn.1001-1714.2005.04.007 URL [本文引用: 1] 摘要
文章对白石砬子山南北坡森林植被垂直分布按20m垂直海拔高度分 段进行了植物种类、分布、多度的详细调查和分析.首次发现了兰科植物双花红门兰(Orchis cyclochila)的分布,填补了辽宁无此植物的空白.同时认为其植被分布有着特殊性,因此用温度随纬度、海拔递减规律估计各种植被分布误差较大.
Forest vegetation vertical distribution in south and north slopes of Baishilazi Mountain [J].https://doi.org/10.3969/j.issn.1001-1714.2005.04.007 URL [本文引用: 1] 摘要
文章对白石砬子山南北坡森林植被垂直分布按20m垂直海拔高度分 段进行了植物种类、分布、多度的详细调查和分析.首次发现了兰科植物双花红门兰(Orchis cyclochila)的分布,填补了辽宁无此植物的空白.同时认为其植被分布有着特殊性,因此用温度随纬度、海拔递减规律估计各种植被分布误差较大.
|
| [10] |
a. 湖北植被地理分布的规律性(上) [J].
湖北植被地理分布的规律性(下)王映明(中国科学院武汉植物研究所武汉430074)2植被的垂直地带性分布规律山地植被的垂直分布,不仅受山体的高低大小、坡向乃至土壤基质等因素的影响,尤其是山地海拔高度所引起的气候因素的变化,直接导致植被从山体的基带向上有...
a. The geographically distributional regularity of the vegetation in Hubei Province (I) [J].
湖北植被地理分布的规律性(下)王映明(中国科学院武汉植物研究所武汉430074)2植被的垂直地带性分布规律山地植被的垂直分布,不仅受山体的高低大小、坡向乃至土壤基质等因素的影响,尤其是山地海拔高度所引起的气候因素的变化,直接导致植被从山体的基带向上有...
|
| [11] |
b. 湖北植被地理分布的规律性(下) [J].
湖北植被地理分布的规律性(下)王映明(中国科学院武汉植物研究所武汉430074)2植被的垂直地带性分布规律山地植被的垂直分布,不仅受山体的高低大小、坡向乃至土壤基质等因素的影响,尤其是山地海拔高度所引起的气候因素的变化,直接导致植被从山体的基带向上有...
a. The regularity of geographical distributions of the vegetation in Hubei Province (Ⅱ) [J].
湖北植被地理分布的规律性(下)王映明(中国科学院武汉植物研究所武汉430074)2植被的垂直地带性分布规律山地植被的垂直分布,不仅受山体的高低大小、坡向乃至土壤基质等因素的影响,尤其是山地海拔高度所引起的气候因素的变化,直接导致植被从山体的基带向上有...
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
云南景东哀牢山山顶苔藓矮林优势种: 倒卵叶石栎 [J].
云南景东哀牢山; 山顶苔藓矮林; 优势种; 倒卵叶石栎
The dominant species of the dwarfing mossy forest on the summit of Mt. Ailao in Jingdong, Yunnan: Lithoc Arpus Pachyphylloides [J].
云南景东哀牢山; 山顶苔藓矮林; 优势种; 倒卵叶石栎
|
| [16] |
中国山顶苔藓矮曲林的分布及其特征 [J].https://doi.org/10.3969/j.issn.1000-5641.2010.04.006 URL 摘要
利用实地调查和相关文献资料,确定山顶苔藓矮曲林在我国的分布情 况,统计分析其植物组成成分和区系地理成分,并计算群落的相似性.结果表明:山顶苔藓矮曲林在我国南方大部分省区呈"岛屿"状分布,最北限位于河南伏牛 山;其分布地点随着我国三级阶梯的地形变化及各山体具体海拔高度不同而不同;该类群落种类组成较为贫乏,共计蕨类植物16种,种子植物251种;其区系特 点是深受热带亚热带区系的影响,又具有温带区系的性质;山顶苔藓矮曲林群落东西部差异较大,东部地区群落相似性较高;群落丰富度总体呈现出西高东低、南高 北低的趋势.鉴于该类群落生境的独特性和分布的特殊性,一方面其生态系统比较脆弱,应加强保护;另一方面应开展广泛、详细的调查,对其进行系统、深入的研 究.
Distribution and characteristics of the mossy dwarf forests in China [J].https://doi.org/10.3969/j.issn.1000-5641.2010.04.006 URL 摘要
利用实地调查和相关文献资料,确定山顶苔藓矮曲林在我国的分布情 况,统计分析其植物组成成分和区系地理成分,并计算群落的相似性.结果表明:山顶苔藓矮曲林在我国南方大部分省区呈"岛屿"状分布,最北限位于河南伏牛 山;其分布地点随着我国三级阶梯的地形变化及各山体具体海拔高度不同而不同;该类群落种类组成较为贫乏,共计蕨类植物16种,种子植物251种;其区系特 点是深受热带亚热带区系的影响,又具有温带区系的性质;山顶苔藓矮曲林群落东西部差异较大,东部地区群落相似性较高;群落丰富度总体呈现出西高东低、南高 北低的趋势.鉴于该类群落生境的独特性和分布的特殊性,一方面其生态系统比较脆弱,应加强保护;另一方面应开展广泛、详细的调查,对其进行系统、深入的研 究.
|
| [17] |
海南岛五指山的森林植被Ⅰ: 五指山的森林植被类型 [J].
本部分仅对五指山森林植被类型进行科学的划分和描述。五指山森林植被自下而上可划分为热带低地雨林(海拔700~1000m)、热带山地雨林(海拔1000~1400m)、热带亚高山矮林(海拔1400~1700m)、热带山顶灌丛(海拔1700~1867m)等4个原生植被类型,共10个群系。
The forest vegetation of Wu-zhi Shan Mountain in Hainan Island I: The forest vegetation types of Wu-zhi Shan Mountain in Hainan Island [J].
本部分仅对五指山森林植被类型进行科学的划分和描述。五指山森林植被自下而上可划分为热带低地雨林(海拔700~1000m)、热带山地雨林(海拔1000~1400m)、热带亚高山矮林(海拔1400~1700m)、热带山顶灌丛(海拔1700~1867m)等4个原生植被类型,共10个群系。
|
| [18] |
海南岛五指山的森林植被II: 五指山森林植被的植物种群分析与森林结构分析 [J].
The forest vegetation of Wu-zhi Shan Mountain in Hainan Island II: The plant population analysis of forest structures analysis of the vegetation Wu-zhi Shan Mountain in Hainan Island [J].
|
| [19] |
中国的猴头杜鹃矮林 [J].URL 摘要
猴头杜鹃矮林大多分布于中国亚热带东部地区的平缓山脊和山顶。其面积虽小,但分布广泛。猴头杜鹃矮林是地形顶极和半地带性的植被类型,它反映着我国亚热带地区海拔1000─2000m地段的大气候状况,但也反映山顶局部气候因素的影响。本文阐述了该林型的一般群落学特点及生态、经济效益。
On the Rhododendron Simiarum elfin forest of China [J].URL 摘要
猴头杜鹃矮林大多分布于中国亚热带东部地区的平缓山脊和山顶。其面积虽小,但分布广泛。猴头杜鹃矮林是地形顶极和半地带性的植被类型,它反映着我国亚热带地区海拔1000─2000m地段的大气候状况,但也反映山顶局部气候因素的影响。本文阐述了该林型的一般群落学特点及生态、经济效益。
|
| [20] |
伏牛山森林群落物种多样性研究Ⅰ: 群落垂直分布与物种丰富度 [J].URL 摘要
系统研究了河南伏牛山国家自然保护区森林群落的垂直分布和组成群落的乔木,灌木,草本的不同种类及其个体数以及在海拔垂直梯度上的变化特征。探讨了不同群落类型与物种丰富度间的关系。
Studies on forest community diversity in Funiushan Mountain I: Vertical distribution of community and species richness [J].URL 摘要
系统研究了河南伏牛山国家自然保护区森林群落的垂直分布和组成群落的乔木,灌木,草本的不同种类及其个体数以及在海拔垂直梯度上的变化特征。探讨了不同群落类型与物种丰富度间的关系。
|
| [21] |
|
| [22] |
广东乳阳八宝山自然保护区的植被特点 [J].URL 摘要
乳阳八宝山自然保护区,地处湘粤交界的南岭山脉东南侧,介于112°56′08″E~113°04′18″E,24°30′28″N~24°48′09″N之间,占地3301.9公顷。地形复杂,峰峦叠嶂,为广东省最高峻雄伟的山地。特殊的地理位置和复杂的自然条件使这里的植被特点复杂而多样。主要表现在: 1.植物区系地理成分复杂。在属的地理成分结构中,除了未见有中亚分布类型外,其它14类成分均有代表,其中又以热带、亚热带成分占绝对优势,但山上亦有一定比重的北温带成分。 2.植被的外貌终年常绿。生活型谱:Ph(83.33%)H(8.34%)Ch(4.76%)Cr(3.57%)T(0%);叶级谱:中叶(57.1%)小叶(29.8%)微叶(17.1%)大叶(6.0%),这与典型常绿阔叶林的叶级谱有较大区别,在叶质叶缘等方面亦有相似的特征,表现它已明显地向南亚热带、热带森林过渡。 3.植被类型多种多样。主要植被类型有:典型常绿阔叶林、亚热带山地常绿阔叶林、亚热带山地常绿落叶阔叶混交林、亚热带常绿针叶林、亚热带常绿针阔叶混交林、亚热带山地常绿针阔叶混交林、亚热带山顶常绿阔叶苔藓矮林。 4.植被的垂直分布规律明显。自下而上见有:典型常绿阔叶林带(海拔700~800米以下),亚热带山地常绿阔叶林带(海拔700或800米至1600或1700米之间)、亚热带山顶常绿阔叶苔藓矮林带(海拔1600或1700米以上)。 5.原生植被保存面积大,珍稀濒危植物丰富。因此,建议有关部门进一步扩大保护区范围,加强调查研究工作,争取早日建成国家级南岭自然保护区。
Vegetation characters of the Ruyang Babao Mountain Nature Reserve, Guangdong Province [J].URL 摘要
乳阳八宝山自然保护区,地处湘粤交界的南岭山脉东南侧,介于112°56′08″E~113°04′18″E,24°30′28″N~24°48′09″N之间,占地3301.9公顷。地形复杂,峰峦叠嶂,为广东省最高峻雄伟的山地。特殊的地理位置和复杂的自然条件使这里的植被特点复杂而多样。主要表现在: 1.植物区系地理成分复杂。在属的地理成分结构中,除了未见有中亚分布类型外,其它14类成分均有代表,其中又以热带、亚热带成分占绝对优势,但山上亦有一定比重的北温带成分。 2.植被的外貌终年常绿。生活型谱:Ph(83.33%)H(8.34%)Ch(4.76%)Cr(3.57%)T(0%);叶级谱:中叶(57.1%)小叶(29.8%)微叶(17.1%)大叶(6.0%),这与典型常绿阔叶林的叶级谱有较大区别,在叶质叶缘等方面亦有相似的特征,表现它已明显地向南亚热带、热带森林过渡。 3.植被类型多种多样。主要植被类型有:典型常绿阔叶林、亚热带山地常绿阔叶林、亚热带山地常绿落叶阔叶混交林、亚热带常绿针叶林、亚热带常绿针阔叶混交林、亚热带山地常绿针阔叶混交林、亚热带山顶常绿阔叶苔藓矮林。 4.植被的垂直分布规律明显。自下而上见有:典型常绿阔叶林带(海拔700~800米以下),亚热带山地常绿阔叶林带(海拔700或800米至1600或1700米之间)、亚热带山顶常绿阔叶苔藓矮林带(海拔1600或1700米以上)。 5.原生植被保存面积大,珍稀濒危植物丰富。因此,建议有关部门进一步扩大保护区范围,加强调查研究工作,争取早日建成国家级南岭自然保护区。
|
| [23] |
我国山地环境的自然特点及开发利用 [J].
正 我国是一个多山的国家,山地(包括丘陵和山原)约占全国土地总面积的三分之二。这些广泛分布的山地,在我国地理环境中起着巨大的作用,并形成了独特的开发利用和改造问题。因此,对我国山地开展深入的科学研究,在地理学理论上和工农业生产上均具有重大意义。
Physical feature of China's mountain environment and economic problem of its utilization [J].
正 我国是一个多山的国家,山地(包括丘陵和山原)约占全国土地总面积的三分之二。这些广泛分布的山地,在我国地理环境中起着巨大的作用,并形成了独特的开发利用和改造问题。因此,对我国山地开展深入的科学研究,在地理学理论上和工农业生产上均具有重大意义。
|
| [24] |
|
| [25] |
略论川西山地的自然垂直带 [J].
川西山地是指四川盆地以西的高山和高原,包括阿坝藏族自治州、甘孜藏族自治州、凉山彝族自治州、渡口市,以及绵阳地区、成都市、雅安地区和乐山地区西部的山地。这片山地的面积约36万平方公里。 以往,对川西山地自然垂直带曾作了不同程度的研究。 近年来,我们对川西山地自然垂直带进行了较大范围的专题考察。现就概略论述一下川西山地自然垂直带的形成条件、主要类型、基本特征及变异规律。
Trial discussion on the vertical natural zone of the mountains in west Sichuan [J].
川西山地是指四川盆地以西的高山和高原,包括阿坝藏族自治州、甘孜藏族自治州、凉山彝族自治州、渡口市,以及绵阳地区、成都市、雅安地区和乐山地区西部的山地。这片山地的面积约36万平方公里。 以往,对川西山地自然垂直带曾作了不同程度的研究。 近年来,我们对川西山地自然垂直带进行了较大范围的专题考察。现就概略论述一下川西山地自然垂直带的形成条件、主要类型、基本特征及变异规律。
|
| [26] |
|
| [27] |
Studies on upper timberline: Morphology and anatomy of Norway spruce (Picea abies) and stone pine (Pinus cembra) needles from various habitat conditions [J]. |
| [28] |
Tree lines and germ plasm-a study in evolutionary limitations [J].https://doi.org/10.1073/pnas.50.5.860 URL 摘要
Not Available
|
| [29] |
Vegetational zonation in the Rocky Mountains [J].https://doi.org/10.1007/BF02872481 URL [本文引用: 1] 摘要
No Abstract available for this article.
|
| [30] |
Vegetation Mitteleuropas mit den Alpen [M].
|
| [31] |
|
| [32] |
Interpretation of the 'Massenerhebung' effect on tropical mountains [J].https://doi.org/10.1038/229044a0 URL PMID: 16059069 摘要
Abstract THREE types of rain forest can generally be recognized on wet tropical mountains: lowland rain forest, lower Montane rain forest and upper Montane rain forest1–3. These forest types can be defined both by distinctive plant associations1 and by the altitudinal limits within which they lie. These limits, however, vary with the type of mountain. On small, isolated mountains and outlying ridges of major ranges, the upper limit of lowland rain forest is about 700–900 m and that of the lower Montane rain forest about 1,200–1,600 m, whereas on the main ridges of major ranges the limits are higher, approximately 1,200–1,500 m and 1,800–2,300 m, respectively4. This phenomenon is known as the ‘Massenerhebung’ effect.
|
| [51] |
Tertiary climates and floristic relationships at high latitudes in the Northern Hemisphere [J].https://doi.org/10.1016/0031-0182(80)90063-2 URL [本文引用: 1] 摘要
During the Paleocene and Eocene, climates were characterized by a low mean annual range of temperature (a maximum of 10–15°C), a moderate to high mean annual temperature (10–20°C), and abundant precipitation; strong broad-leaved evergreen vegetation extended to almost lat. 60°N during the Paleocene and to well above 61°N during the Eocene. Poleward of the broad-leaved evergreen forests were forests that were broad-leaved deciduous; these deciduous forests, however, were unlike extant broad-leaved deciduous forests in general floristic composition and physiognomy. Coniferous forests probably occupied the northernmost latitudes. At the end of the Eocene, a major climatic deterioration resulted in a high (> 30°C) mean annual range of temperature and a low mean annual temperature (< 10°C). Vegetation represented temperate broad-leaved deciduous and coniferous forests. The Oligocene and Neogene climatic trends represent a decrease in both mean annual range of temperature and mean annual temperature. Tundra vegetation did not appear until late in the Neogene. The present distribution of broad-leaved evergreens concomitant with the principles of plant physiology indicates that present winter light conditions at high latitudes could not support broad-leaved evergreen forest. A possible solution to the problem is to increase winter light by lessening the inclination of the earth's rotational axis.
|

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}