
冬季非均匀增温与积雪及光周期变化对温带植被物候影响研究进展
|
王笑影(1974— ),女,辽宁沈阳人,副研究员,研究方向为生态气象与气候变化。E-mail: wangxy@iaesy.cn |
收稿日期: 2023-12-04
修回日期: 2024-03-11
网络出版日期: 2024-08-22
基金资助
国家重点研发计划项目(2022YFF0801301-1)
中国气象局沈阳大气环境研究所和辽宁省农业气象灾害重点实验室联合开放基金项目(2023SYIAEKFZD05)
Progress of research on the impact of winter non-uniform warming, snow cover, and photoperiod changes on the phenology of temperate vegetation
Received date: 2023-12-04
Revised date: 2024-03-11
Online published: 2024-08-22
Supported by
National Key Research and Development Program(2022YFF0801301-1)
Joint Open Fund of Shenyang Institute of Atmospheric Environment, China Meteorological Administration and Key Laboratory of Agro-meteorological Disasters of Liaoning Province(2023SYIAEKFZD05)
气候变化速率在日尺度和季节尺度上表现出差异性,并且呈现出非均匀增温的特点。特别是在北半球中高纬度地区,夜间的升温速度超过白天,而冬季的升温速度超过夏季。准确评估非均匀增温对陆地生态系统结构和功能的影响,是全球变化研究领域一项重大挑战。论文通过文献综述,系统地分析了冬季非均匀增温、积雪变化以及光周期对春季植被物候特征的影响,同时探讨了这种影响存在的物种特异性、地理位置异质性以及季节补偿效应。研究认为,当前对非均匀增温影响的研究在观测实验能力方面存在明显不足,对光照、温度和水分耦合作用的理解和认识尚不充分,且模型模拟能力有待提高。因此,未来研究应重视开发新型观测和实验方法,例如精心设计控制实验,以深入探究温带地区光照、温度和水分对植物物候的潜在调控机制。这将有助于加强对非均匀增温影响机制的认识和理解,并将这些影响因素纳入模型模拟中,从而提升对季节性气候变化如何塑造植被动态过程的认知和理解。
王笑影 , 周玉科 , 李荣平 , 贾庆宇 , 蔡福 , 温日红 , 谢艳兵 , 赵先丽 , 邹旭东 , 汪宏宇 . 冬季非均匀增温与积雪及光周期变化对温带植被物候影响研究进展[J]. 地理科学进展, 2024 , 43(8) : 1666 -1680 . DOI: 10.18306/dlkxjz.2024.08.015
The rate of climate change exhibits differences at daily and seasonal scales and is characterized by non-uniform warming. Particularly in the mid to high latitudes of the Northern Hemisphere, the rate of warming at night surpasses that during the day, and the rate of warming in winter exceeds that in summer. Accurately assessing the impact of non-uniform warming on the structure and function of terrestrial ecosystems represents a significant challenge in the field of global change research. This article, through a literature review, systematically analyzed the effects of non-uniform warming in winter, changes in snow metrics, and variations in photoperiod on the phenological characteristics of spring vegetation. It also discussed factors such as species specificity, geographic location heterogeneity, and seasonal compensation effects. The research indicated that seasonal changes in temperate regions have a significant impact on vegetation phenological cycle, but existing studies have shortcomings in observation experiments, mechanism understanding, and model simulation. Although controlled experiments are helpful for studying the effects of meteorological factors on vegetation, they may not fully reflect the natural conditions. Remote sensing monitoring provides a macroscopic perspective, but its data accuracy is limited by many factors, and it is difficult to capture subtle phenological changes. Ground observations provide valuable information for climate science, but the distribution of observation sites is sparse and the data uncertainty is large. In terms of the mechanism of impact, the interactions of light, temperature, and water and their effects on phenology are not fully understood, and the carry-over lag effect between seasons is not well understood either. In terms of model simulation, it is difficult to parameterize the plant phenology model, the study of parameter thresholds is insufficient, and the influence of other biotic and abiotic factors is not fully considered. These challenges limit the accuracy of predictions of plant phenological changes. Therefore, future research is needed to develop novel observational experimental methods to accurately distinguish plant responses under different environmental conditions and to validate model predictions under more natural conditions. This includes improving warming experiments, accounting for the effects of nighttime warming, and encouraging research based on global field observations. It is also necessary to strengthen the understanding of the mechanism of influence of non-uniform warming, especially the light, heat, and water requirements of spring phenology, and the complex effects of winter and spring warming on leaf phenology. In addition, research should focus on fall phenological processes and the risk of late spring frost. In terms of modeling, non-uniform warming and its associated impacts need to be incorporated into models to improve the ability to predict the response of temperate ecosystems to climate change. This includes precise measurements of the onset of ecological dormanism in trees, combined with physiological studies, and consideration of seasonal climate change effects on carbon sequestration and cycling in terrestrial ecosystem models. At the same time, the impact of seasonal snowfall should be considered and the understanding of photoperiodic effects should be deepened to assess the potential impacts of climate warming on terrestrial ecosystems, thereby enhancing our comprehension of the impacts of climate change on vegetation at the seasonal scale.
Key words: non-uniform warming; snow cover; photoperiod; winter; vegetation phenology
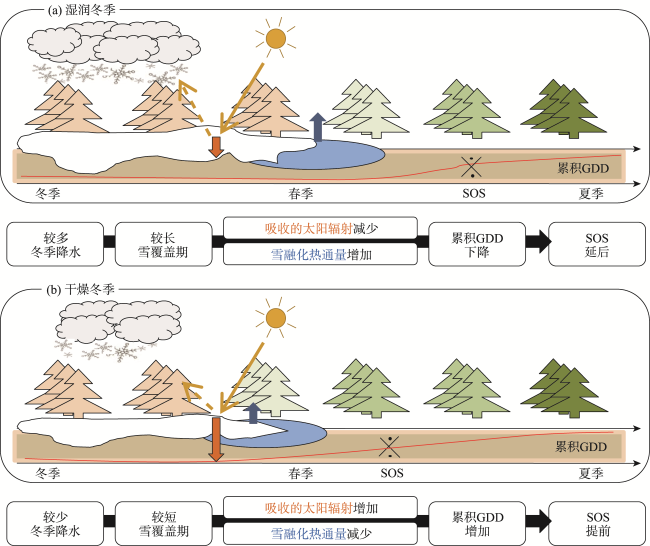
图1 冬季降雪通过改变春季平均吸收太阳辐射(橙色箭头)、融雪热通量(蓝色箭头)和累积积温(GDD,红线)对春季物候期(SOS)的影响机制示意图注:根据参考文献[49]进行修改。 Fig.1 Schematic diagram of the mechanism of influence of winter snowfall on spring phenology (SOS) by changing spring average absorbed solar radiation (orange arrow), snowmelt heat flux (blue arrow), and cumulative accumulated temperature (GDD, red line) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
承继成, 郭华东, 史文中, 等. 遥感数据的不确定性问题[M]. 北京: 科学出版社, 2004.
[
|
| [71] |
|
| [72] |
周玉科. 基于数码照片的植被物候提取多方法比较研究[J]. 地理科学进展, 2018, 37(8): 1031-1044.
[
|
| [73] |
范德芹, 赵学胜, 朱文泉, 等. 植物物候遥感监测精度影响因素研究综述[J]. 地理科学进展, 2016, 35(3): 304-319.
[
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
刘正佳. 气象数据的不确定性对 CEVSA 模型模拟结果的影响研究[D]. 济南: 山东师范大学, 2012.
[
|
| [78] |
沈军. 气象自动观测站数据处理方法研究[D]. 长沙: 中南大学, 2011.
[
|
| [79] |
段晚锁, 汪叶, 霍振华, 等. 数值天气预报和气候预测的集合预报方法: 思考与展望[J]. 气候与环境研究, 2019, 24(3): 396-406.
[
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}