
植物始花期对气候变化响应的激素调控机理研究进展
张湜溪(1990— ),女,博士后,研究领域为环境分析化学。E-mail: zhangsx@igsnrr.ac.cn
收稿日期: 2018-09-05
要求修回日期: 2019-03-22
网络出版日期: 2019-07-28
基金资助
中国科学院战略性先导科技专项(A类)(XDA19040301)
国家重点研发计划项目(2018YFA0606102)
国家自然科学基金项目(41427805)
Progress of responses of first flowering date to climate change and the correlations of plant hormone regulation
Received date: 2018-09-05
Request revised date: 2019-03-22
Online published: 2019-07-28
Supported by
Strategic Priority Research Program of Chinese Academy of Sciences (Class A), No. XDA19040301
National Key Research and Development Program of China, No. 2018YFA0606102
National Natural Science Foundation of China, No. 41427805.
Copyright
植物物候对气候变化的响应非常敏感,是指示全球变化对生态系统影响的重要证据。其中植物花期变化影响植物繁殖与进化,具有重要意义。大量研究表明,随着全球气候变暖,北半球植物始花期普遍提前。而关于气候变暖对植物始花期影响的内在机理并没有明确解释。植物激素是植物体内对植物开花等生理活动有显著调控作用的有机物,可对环境刺激作出响应并直接参与调控植物始花期,导致始花期的提前或延后。对植物激素在植物中表达与变化的精确测定有助于了解植物始花期对气候变化响应的内在机理。论文综述了植物激素对植物开花时间的调控作用以及目前植物激素最先进的检测方法,探讨了植物激素及相关代谢产物在植物体内的含量变化对植物始花期的影响;提出通过植物激素研究植物始花期对气候变化响应的激素调控机理,为探索植物物候对气候变化响应提供新的研究思路与手段。
张湜溪 , 戴君虎 , 葛全胜 . 植物始花期对气候变化响应的激素调控机理研究进展[J]. 地理科学进展, 2019 , 38(7) : 1045 -1055 . DOI: 10.18306/dlkxjz.2019.07.009
Plant phenology is an important indicator of the impact of global change on biological systems because it is very sensitive to climate change. Flowering phenology is of great importance since it represents the major developmental transition of plants from the vegetative to the reproductive stage. Many existing studies have identified the advancement in first flowering date in response to global warming. However, the mechanism of first flowering date dependency on climate change remains obscure. Plant hormones play pivotal roles in almost every aspect in plant life including flowering. To understand the regulation of plant flowering phenology in response to climate change by plant hormones, accurate and efficient measurements of plant hormones are required. In this review, the functions and possible links between plant hormones and the change in first flowering date were discussed. Also, recently developed methods for plant hormone detection were briefly summarized, providing scientific techniques for understanding the underlying mechanisms of plant phenology in response to environmental stimuli.
Key words: phenology; first flowering date; plant hormone; mass spectrometry
Tab.1 Chemical structures of auxins, jasmonic acid, salicylic acid, cytokinins, gibberellins, abscisic acid and related compounds表1 几种主要植物激素类型和结构 |
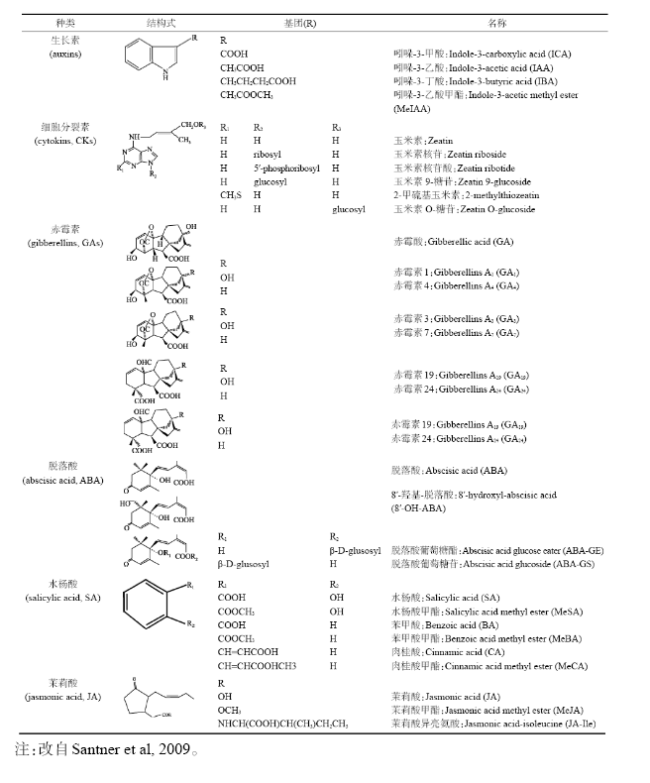
Fig.1 The molecular mechanism suggested for Flowering Locus C (FLC) expression图1 赤霉素(GA)介导的DELLA蛋白调控Flowering Locus C (FLC)表达机理 |
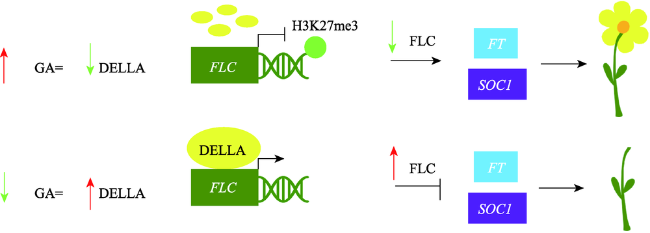
Fig.2 The participation of salicylic acid (SA) in SUMO modification of Flowering Locus D (FLD)图2 水杨酸(SA)介导的Flowering Locus D(FLD)类泛素化修饰调控路径 |
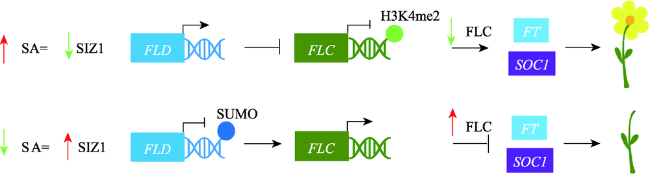
Fig.3 Multiple hormones regulating flowering time图3 多种植物激素调控植物开花信号传导通路 |
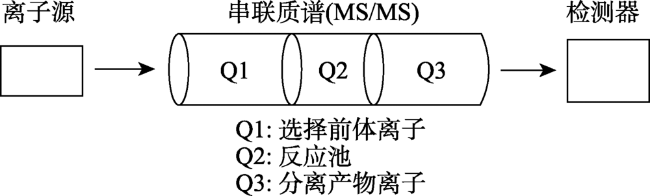
Fig.4 Flow chart of the strategy for plant hormone analysis by MS/MS图4 串联质谱(MS/MS)检测原理示意图 |
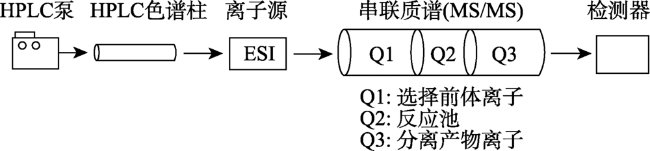
Fig.5 Flow chart of the strategy for plant hormone analysis by HPLC-ESI-MS/MS图5 HPLC-ESI-MS/MS检测植物激素原理示意图 |
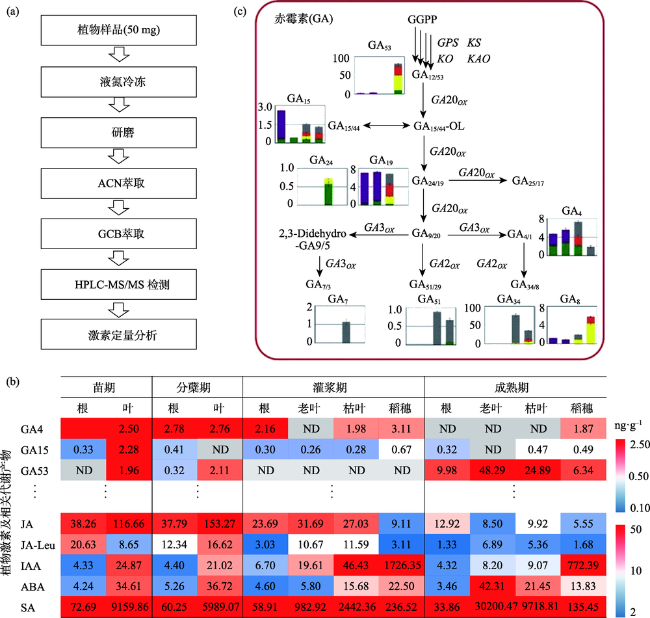
Fig.6 A rapid investigation approach for spatiotemporal distribution of phytohormones in rice by UPLC-electrospray ionization-tandem mass spectrometry (UPLC-ESI-MS/MS)图6 基于液相色谱-质谱联用(UPLC-ESI-MS/MS)的植物激素在水稻不同植物组织的快速检测方法 |
The authors have declared that no competing interests exist.
| [1] |
[
|
| [2] |
[
|
| [3] |
[
|
| [4] |
[
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}