Progress and prospects of research on shells of Radix sp. in lakes on the Tibetan Plateau
CHEN Feng1, 2, , FENG Jinliang3
1. Department of Environmental Economics, Shanxi University of Finance and Economics, Taiyuan 030006, China2. Institute of Surface Processes and Ecological Environment, Shanxi University of Finance and Economics, Taiyuan 030006, China3. Institute of Tibetan Plateau Research, CAS, Beijing 100101, China
Palaeohydrology and palaeohydrochemical reconstruction have always been one of the most challenging issues in lake research, and one of the main reasons for this is the lack of suitable archives. Radix sp. fossil shells are widely distributed in lakes, alluvial-lacustrine successions, and high level lake sediments on the Tibetan Plateau. These biogenic carbonate shells are a kind of promising high-resolution carrier of the environmental information. For example, the δ13Cshell values of the Radix shells are controlled by the δ13C of lake water dissolved inorganic carbon; the δ18Oshell values provide useful information about the isotopic composition of the ambient waters; and the Sr/Ca ratio of the Radix shells is significantly positively correlated with both the Sr/Ca ratio and the conductivity of the lake water. Therefore, the δ13Cshell and δ18Oshell values and element of the Radix sp. shells have been gradually used to provide the palaeohydrological and palaeoclimatic information on the Tibetan Plateau and other regions in existing studies. In addition, the 87Sr/86Srshell of Radix sp. reflects significantly the 87Sr/86Sr of the water in which the shells were formed. However, there are still many scientific issues to be explored: it is unclear how the shells of Radix sp. record the hydrological, hydrochemical, and other information of its ambient water, and how we reconstruct palaeoenvironment information based on the fossil shells of Radix sp. As a new environmental information archive, Radix sp. and their following aspects in existing research on lakes of the Tibetan Plateau were reviewed in this study, including: the classification systems of Radix sp., habitat studies of Radix sp. and applications, proxies of Radix sp. shell (δ13Cshell, δ18Oshell, 87Sr/86Sr, Sr/Ca and Mg/Ca), and palaeohydrochemical reconstruction in lakes, in order to explore the prospects for future study on Radix sp. This review found that significant unanswered questions still remain. For example, the influence of lake water chemistry and water depth on the distribution and survival of Radix sp. is unclear; it is not clear whether inter-species differences have an impact on the results of lake sediment and palaeoenvironmental studies; species level classification of Radix sp., oxygen isotope balance, and trace element need further study; and more environmental records of Radix sp. shells are needed, among others.
Keywords:plateau lakes
;
Radix sp.
;
stable carbon and oxygen isotopes
;
strontium isotope
;
shell element ratio
;
palaeohydrochemical reconstruction
CHENFeng, FENGJinliang. Progress and prospects of research on shells of Radix sp. in lakes on the Tibetan Plateau[J]. Progress in Geography, 2018, 37(10): 1430-1441 https://doi.org/10.18306/dlkxjz.2018.10.012
萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986)。萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012)。萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998)。因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行。值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990)。尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012)。因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息。例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000)。Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论。
萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存。Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动。因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975)。同时,这种习性在古环境研究中也得到了验证。Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期。值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979)。总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m。Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养。萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998)。因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977)。另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上。
长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016)。近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017)。其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制。此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息。在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化。由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体。
近年来,Chen等(2016)建立了青藏高原中南部区域上多个湖泊中现生萝卜螺属壳体地球化学与其宿生水体水化学之间的关系,即萝卜螺属壳体δ18Oshell的变化主要反映了湖水氧同位素(δ18Owater)的变化,例如湖水蒸发或者降水δ18O的变化(Tyler et al, 2008);萝卜螺属壳体δ13Cshell取决于湖水的溶解无机碳(DIC)的δ13CDIC,并初步探讨了萝卜螺属壳体—水体δ18O分馏系数(变化范围在1.031~1.040)及其温度依赖性(较低)。在此基础上,Chen等(2017)利用纳木错周边的高湖面沉积物中的萝卜螺属壳体化石的δ18Oshell和δ13Cshell结合高湖面沉积物的理化性质探讨了纳木错中晚全新世高湖面的变化机制。
在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016)。水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017)。大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响。但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013)。因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放)。河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016)。可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低。基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态。
现生萝卜螺属壳体主要包含三种元素,即Ca、Sr和Mg,其含量的变化范围分别为:Ca 35.8~39.67(%)、Sr 849.5~2383(ppm)、Mg 24.35~164.6(ppm)(Chen et al, 2016)。上述数据表明,萝卜螺属壳体钙的含量很高,基本上是纯净的碳酸盐;不同湖泊获取的萝卜螺属壳体的特征元素具有较大的差异性,这与其所生活的水体环境有关(Chen et al, 2016)。
The application of Mg/Ca, Sr/Ca and 87Sr/86Sr ratios of biogenic shells in reconstruction of palaeoclimate and palaeoenvironment as examplified by Xiaodukou section
Carbonate deposition, Pyramid Lake subbasin, Nevada: 3. The use of 87Sr values in carbonate deposits (tufas) to determine the hydrologic state of paleolake systems
Potential forcing mechanisms of Holocene lake-level changes at Nam Co, Tibetan Plateau: Inferred from the stable isotopic composition of shells of the gastropod Radix
Ostracod Mg/Sr/Ca and 87Sr/86 Sr geochemistry from Tibetan lake sediments: Implications for early to mid-Pleistocene Indian monsoon and catchment weathering
Early to mid-Holocene lake high-stand sediments at Lake Donggi Cona, northeastern Tibetan Plateau, China-comment to the paper published by Dietze et al., Quaternary Research 79(2013), 325-336
Stable isotope composition of the Holsteinian (MIS 11) freshwater snail Valvata piscinalis (O. F. Müller, 1774) from eastern Poland and its palaeoenvironmental implications
[J]. Journal of Quaternary Science, 31(4): 622-630.
Sclerochronological oxygen and carbon isotope ratios in Radix (Gastropoda) shells indicate changes of glacial meltwater flux and temperature since 4200 cal yr BP at Lake Karakul, eastern Pamirs (Tajikistan)
Sub-seasonal oxygen and carbon isotope variations in shells of modern Radix sp. (Gastropoda) from the Tibetan Plateau: Potential of a new archive for palaeoclimatic studies
Oxygen and carbon isotope patterns archived in shells of the aquatic gastropod Radix: Hydrologic and climatic signals across the Tibetan Plateau in sub-monthly resolution
Chemical and isotopic (O, H, C) composition of surface waters in the catchment of Lake Donggi Cona (NW China) and implications for paleoenvironmental reconstructions
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... ).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... )、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... )、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... )和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... 现生萝卜螺属壳体主要包含三种元素,即Ca、Sr和Mg,其含量的变化范围分别为:Ca 35.8~39.67(%)、Sr 849.5~2383(ppm)、Mg 24.35~164.6(ppm)(Chen et al, 2016).上述数据表明,萝卜螺属壳体钙的含量很高,基本上是纯净的碳酸盐;不同湖泊获取的萝卜螺属壳体的特征元素具有较大的差异性,这与其所生活的水体环境有关(Chen et al, 2016). ...
... ).上述数据表明,萝卜螺属壳体钙的含量很高,基本上是纯净的碳酸盐;不同湖泊获取的萝卜螺属壳体的特征元素具有较大的差异性,这与其所生活的水体环境有关(Chen et al, 2016). ...
... Relationship between Mg/Ca molar ratio of Radix sp. shells and hydrochemistry. Data from Chen et al (2016) ...
藏中南典型湖泊全新世高湖面沉积及其古环境意义
12
2016
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... ).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... )、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... )、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... )和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... 现生萝卜螺属壳体主要包含三种元素,即Ca、Sr和Mg,其含量的变化范围分别为:Ca 35.8~39.67(%)、Sr 849.5~2383(ppm)、Mg 24.35~164.6(ppm)(Chen et al, 2016).上述数据表明,萝卜螺属壳体钙的含量很高,基本上是纯净的碳酸盐;不同湖泊获取的萝卜螺属壳体的特征元素具有较大的差异性,这与其所生活的水体环境有关(Chen et al, 2016). ...
... ).上述数据表明,萝卜螺属壳体钙的含量很高,基本上是纯净的碳酸盐;不同湖泊获取的萝卜螺属壳体的特征元素具有较大的差异性,这与其所生活的水体环境有关(Chen et al, 2016). ...
... Relationship between Mg/Ca molar ratio of Radix sp. shells and hydrochemistry. Data from Chen et al (2016) ...
纳木错高湖面时期的溢流:基于萝卜螺壳体氧同位素及地貌证据
7
2017
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... ; 陈锋等, 2017). ...
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
... ; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... ; 陈锋等, 2017). ...
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
... ; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
新时代自然地理学发展的思考
1
2018
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
西藏札达盆地腹足类组合及生物地层学
2
2012
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
西藏札达盆地腹足类组合及生物地层学
2
2012
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
全新世大暖期时中国夏季风向西北大幅度漂移的一个古生物地层学证据
1
2010
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
柴达木盆地察尔汗古湖贝壳堤剖面碳酸盐和瓣鳃化石碳氧稳定同位素
1
2007
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
柴达木盆地察尔汗古湖贝壳堤剖面碳酸盐和瓣鳃化石碳氧稳定同位素
1
2007
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
青藏高原大湖期
1
2000
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
青藏高原大湖期
1
2000
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
试论西藏全新世古地理的演变
2
1982
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
试论西藏全新世古地理的演变
2
1982
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
淮河水系耳萝卜螺生态的初步研究
2
1998
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
淮河水系耳萝卜螺生态的初步研究
2
1998
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
太湖椭圆萝卜螺的食物来源分析
1
2008
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
太湖椭圆萝卜螺的食物来源分析
1
2008
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
淡水软体动物壳化石碳氧同位素组成及其在恢复古气候环境中的应用
1
1990
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
淡水软体动物壳化石碳氧同位素组成及其在恢复古气候环境中的应用
1
1990
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
基于MODIS的青藏高原湖泊透明度遥感反演
1
2017
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
基于MODIS的青藏高原湖泊透明度遥感反演
1
2017
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
西藏尼洋河水生生物群落时空动态及与环境因子关系: 3. 大型底栖动物
2
2014
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
西藏尼洋河水生生物群落时空动态及与环境因子关系: 3. 大型底栖动物
2
2014
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
锶同位素在沉积学中的研究与进展
1
2007
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
锶同位素在沉积学中的研究与进展
1
2007
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
7
1979
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
7
1979
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
耳萝卜螺Radix auricularia(L. 1758)的形态结构
1
1963
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
青藏高原东部兴措湖生物壳体元素及同位素记录的气候环境信息
1
2001
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
青藏高原东部兴措湖生物壳体元素及同位素记录的气候环境信息
1
2001
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
锶同位素在水文地质学研究中的应用进展
1
2009
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
锶同位素在水文地质学研究中的应用进展
1
2009
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
藏南沉错钻孔硅藻组合与湖水古盐度定量恢复
1
2003
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
藏南沉错钻孔硅藻组合与湖水古盐度定量恢复
1
2003
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
水文学研究进展与展望
1
2018
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
水文学研究进展与展望
1
2018
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
湖泊沉积记录的古洪水事件识别及与气候关系
1
2015
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
湖泊沉积记录的古洪水事件识别及与气候关系
1
2015
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
长江中下游地区典型湖泊摇蚊亚化石-湖水总磷定量模型研究
1
2006
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
长江中下游地区典型湖泊摇蚊亚化石-湖水总磷定量模型研究
1
2006
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
西藏纳木错扎弄淌剖面 10000 年以来的沉积间断和环境变化记录
1
2003
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
西藏纳木错扎弄淌剖面 10000 年以来的沉积间断和环境变化记录
1
2003
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
Carbonate deposition, Pyramid Lake subbasin, Nevada: 3. The use of 87Sr values in carbonate deposits (tufas) to determine the hydrologic state of paleolake systems
1
1996
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
East Asian summer monsoon precipitation variability since the last deglaciation
1
2015
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
Relationship between the shell geochemistry of the modern aquatic gastropod Radix and water chemistry of lakes of the Tibetan Plateau
... 现生萝卜螺属壳体Mg/Ca摩尔比与水化学之间的关系 (数据源自Chen et al, 2016) ...
Potential forcing mechanisms of Holocene lake-level changes at Nam Co, Tibetan Plateau: Inferred from the stable isotopic composition of shells of the gastropod Radix
1
2017
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
Seawater Sr isotope variation over the past 300 kyr and influence of global climate cycles
1
1992
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
Environmental control of strontium and magnesium in Mytilus
1
1965
... 前人研究已表明,软体动物壳体的Sr/Ca比值与其生活水域的水体的温度(Dodd, 1965; Surge et al, 2006)或者盐度(Wolf et al, 1967; Eisma et al, 1976)存在显著的关系.Chen等(2016)对青藏高原中南部湖泊中现生萝卜螺属壳体元素与其宿生水体元素之间的关系进行了探讨,结果表明:萝卜螺属壳体的Sr/Cashell与其生活水体的Sr/Cawater呈显著的线性正相关(Sr/Cashell=0.118×Sr/Cawater+0.0008; R2=0.78);Sr/Cashell与湖水的电导率(EC)也呈显著的线性正相关(Sr/Cashell=9×10-7×EC+0.0001; R2=0.71);Sr/Cashell与其生活水体的温度和pH值均没有明显的关系;现生萝卜螺属壳体Sr的分配系数的变化范围在0.167~0.867之间,平均值是0.317,这一分配系数与以往的研究结果相接近.基于萝卜螺属壳体与宿生水体已建立的地球化学关系,陈锋等(2017)结合纳木错高湖面沉积物中萝卜螺属壳体化石定量重建了该湖4.4-2.2 ka期间湖水的古Sr/CaPalaeo-water和古电导率. ...
Shell characteristics, isotopic composition and trace-element contents of some euryhaline molluscs as indicators of salinity
1
1976
... 前人研究已表明,软体动物壳体的Sr/Ca比值与其生活水域的水体的温度(Dodd, 1965; Surge et al, 2006)或者盐度(Wolf et al, 1967; Eisma et al, 1976)存在显著的关系.Chen等(2016)对青藏高原中南部湖泊中现生萝卜螺属壳体元素与其宿生水体元素之间的关系进行了探讨,结果表明:萝卜螺属壳体的Sr/Cashell与其生活水体的Sr/Cawater呈显著的线性正相关(Sr/Cashell=0.118×Sr/Cawater+0.0008; R2=0.78);Sr/Cashell与湖水的电导率(EC)也呈显著的线性正相关(Sr/Cashell=9×10-7×EC+0.0001; R2=0.71);Sr/Cashell与其生活水体的温度和pH值均没有明显的关系;现生萝卜螺属壳体Sr的分配系数的变化范围在0.167~0.867之间,平均值是0.317,这一分配系数与以往的研究结果相接近.基于萝卜螺属壳体与宿生水体已建立的地球化学关系,陈锋等(2017)结合纳木错高湖面沉积物中萝卜螺属壳体化石定量重建了该湖4.4-2.2 ka期间湖水的古Sr/CaPalaeo-water和古电导率. ...
Constraining water sources and hydrologic processes from the isotopic analysis of water and dissolved strontium, Lake Junin, Peru
1
2005
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
Life cycle of Lymnaea peregra (Gastropoda: Pulmonata) in the Leicester canal, UK, with an estimate of annual production
2
1986
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
Application of strontium isotopes to understanding the hydrology and paleohydrology of the Altiplano, Bolivia-Peru
1
2003
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
δ18O in mollusk shells from Pliocene Lake Hadar and modern Ethiopian Lakes: Implications for history of the Ethiopian monsoon
1
2002
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
The 87Sr/86Sr ratios of lacustrine carbonates and lake-level history of the Bonneville paleolake system
2
2004
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
... ; Hart et al, 2004). ...
Large Holocene summer temperature oscillations and impact on the peopling of the northeastern Tibetan Plateau
1
2016
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
δ18O and δ13C in fossil shells of Radix sp. from the sediment succession of a dammed palaeo-lake in the Yarlung Tsangpo Valley, Tibet, China
6
2017
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... 与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... 值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
... )、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
Lake level reconstruction for 12.8-2.3 ka of the Ngangla Ring Tso closed-basin lake system, southwest Tibetan Plateau
1
2015
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
High late Miocene-Pliocene elevation of the Zhada Basin, southwestern Tibetan Plateau, from carbonate clumped isotope thermometry
1
2015
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
Radix spp.: Identification of trematode intermediate hosts in the Czech Republic
1
2012
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
Ostracod Mg/Sr/Ca and 87Sr/86 Sr geochemistry from Tibetan lake sediments: Implications for early to mid-Pleistocene Indian monsoon and catchment weathering
1
2011
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
The freshwater snails (mollusca: gastropoda) of New York State
1
1992
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
Molecular diversity of Trichobilharzia franki in two intermediate hosts (Radix auricularia and Radix peregra): A complex of species
1
2010
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
Permian strontium isotope stratigraphy
1
2018
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
Palaeoclimate interpretation of stable isotope data from lake sediment archives
1
2004
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
Alkenone-based reconstruction of late-Holocene surface temperature and salinity changes in Lake Qinghai, China
1
2006
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
Modern pollen distributions in Qinghai-Tibetan Plateau and the development of transfer functions for reconstructing Holocene environmental changes
1
2011
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
Quantitative reconstruction of lake conductivity in the Quaternary of the Near East (Israel) using ostracods
2
2010
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
Quantitative relationship between water depth and sub-fossil ostracod assemblages in Lake Donggi Cona, Qinghai Province, China
2
2010
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
Mid to Late Holocene palaeoenvironment of Lake Eastern Juyanze (north-western China) based on ostracods and stable isotopes
2
2002
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
Spatial variability of 14C reservoir effects in Tibetan Plateau lakes
1
2013
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
Early to mid-Holocene lake high-stand sediments at Lake Donggi Cona, northeastern Tibetan Plateau, China-comment to the paper published by Dietze et al., Quaternary Research 79(2013), 325-336
1
2015
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
Lakes and snails: Environment and Gastropoda in 1500 Norwegian lakes, ponds and rivers
1
1990
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
Isotopic tracers of paleohydrologic change in large lakes of the Bolivian Altiplano
2
2011
... 陆地与海洋锶等同位素的研究,是目前全球变化以及古生态环境研究的前沿之一.一个很重要的原因是由于锶同位素与碳、氧和硫这样的轻同位素不同,相变、蒸发作用、化学状态或生物同化作用对Sr同位素分馏影响不明显.锶同位素被广泛应用于海平面变化、湖面变化、气候变化、风化作用、地层年代学与地层学对比、水岩作用以及地下水的循环与演化研究(Dia et al, 1992; 刘昊年等, 2007; Jin et al, 2011; Placzek et al, 2011; Korte et al, 2018).此外,在沉积环境中的87Sr/86Sr比值仅有的变化是由于不同来源Sr的混合造成的(许翔等, 2009).因此,87Sr/86Sr在研究物质迁移和变化过程中是有效的示踪剂,可用于追踪海面或者湖面的变化机制.其理论依据为:在地质历史进程中,海水的87Sr/86Sr值是随时间变化的函数,地质年代越久远,87Sr积累就越多,87Sr/86Sr值也就越高.因此,可根据沉积物87Sr/86Sr组成及其变化规律判读其物质来源.湖泊沉积物中存在大量的腹足纲生物壳体(方解石和文石壳体),这些壳体记录了形成时水体的87Sr/86Sr值.利用这类壳体记录的锶同位素进行示踪,已成功地应用于古湖泊的水文和气候演化等研究(Grove et al, 2003; Hart et al, 2004; Flusche et al, 2005; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
... ; Placzek et al, 2011),如在追踪湖面的变化机制方面已有一些成果(Benson et al, 1996; Hart et al, 2004). ...
Geochemical variation in microstructural shell layers of the southern quahog (Mercenaria campechiensis): Implications for reconstructing seasonality
1
2006
... 前人研究已表明,软体动物壳体的Sr/Ca比值与其生活水域的水体的温度(Dodd, 1965; Surge et al, 2006)或者盐度(Wolf et al, 1967; Eisma et al, 1976)存在显著的关系.Chen等(2016)对青藏高原中南部湖泊中现生萝卜螺属壳体元素与其宿生水体元素之间的关系进行了探讨,结果表明:萝卜螺属壳体的Sr/Cashell与其生活水体的Sr/Cawater呈显著的线性正相关(Sr/Cashell=0.118×Sr/Cawater+0.0008; R2=0.78);Sr/Cashell与湖水的电导率(EC)也呈显著的线性正相关(Sr/Cashell=9×10-7×EC+0.0001; R2=0.71);Sr/Cashell与其生活水体的温度和pH值均没有明显的关系;现生萝卜螺属壳体Sr的分配系数的变化范围在0.167~0.867之间,平均值是0.317,这一分配系数与以往的研究结果相接近.基于萝卜螺属壳体与宿生水体已建立的地球化学关系,陈锋等(2017)结合纳木错高湖面沉积物中萝卜螺属壳体化石定量重建了该湖4.4-2.2 ka期间湖水的古Sr/CaPalaeo-water和古电导率. ...
Stable isotope composition of the Holsteinian (MIS 11) freshwater snail Valvata piscinalis (O. F. Müller, 1774) from eastern Poland and its palaeoenvironmental implications
1
2016
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
Sclerochronological oxygen and carbon isotope ratios in Radix (Gastropoda) shells indicate changes of glacial meltwater flux and temperature since 4200 cal yr BP at Lake Karakul, eastern Pamirs (Tajikistan)
3
2014
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 长期以来,湖泊中腹足类生物碳酸盐壳体的稳定碳氧同位素及元素被广泛用于推测环境和气候的变化(李玉成等, 1990; 吴敬禄等, 2001; Leng et al, 2004; 常凤琴等, 2007; Szymanek, 2016).近年,萝卜螺属壳体的稳定碳氧同位素(δ13Cshell、δ18Oshell)及元素已开始被用于了解青藏高原及其他地区的古水文、古水化学和古气候的信息(Hailemichael et al, 2002; Mischke et al, 2002; Taft et al, 2014; Chen et al, 2017; 陈锋等, 2017).其中,根据现生萝卜螺属的捕获时间及其生长规律,Taft等(2012)和Taft等(2013)对青藏高原9个湖泊的现生萝卜螺属按壳体的生长线连续测试稳定碳氧同位素,并同湖区的气象数据对比发现,在季节到逐月的时间尺度上,萝卜螺属壳体的δ13C和δ18O基本上反映的是水体δ13C和δ18O变化,进而首次阐述了现生萝卜螺属壳体能在半个月分辨率的水平上记录青藏高原的水文和气候信息;随后还讨论了影响萝卜螺属壳体δ13Cshell、δ18Oshell的可能机制.此外,鉴于萝卜螺属壳体化石常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中,因而认为萝卜螺属壳体是记录青藏高原上古季风强度变化的一种极有潜力的载体,利用萝卜螺属壳体化石记录可以重建几千万年前的年际或季节变化环境信息.在此基础上,Taft等(2014)利用萝卜螺属壳体化石的δ13Cshell、δ18Oshell结合介形虫壳体和湖泊自生文石δ18O,重建了帕米尔高原东部的卡拉库里湖4200 cal yr BP 以来的冰川融水和温度变化的情况;研究结果同时表明,萝卜螺属壳体化石的δ18Oshell同介形虫壳体δ18O在长时间尺度上保持了一致的变化趋势,并指示了降水同位素组成的变化.由此进一步表明,萝卜螺属壳体化石同介形虫壳体化石环境记录一样,也可作为一种重要的可信的环境记录载体. ...
Sub-seasonal oxygen and carbon isotope variations in shells of modern Radix sp. (Gastropoda) from the Tibetan Plateau: Potential of a new archive for palaeoclimatic studies
6
2012
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
... 在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
Oxygen and carbon isotope patterns archived in shells of the aquatic gastropod Radix: Hydrologic and climatic signals across the Tibetan Plateau in sub-monthly resolution
3
2013
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
Tibetan Plateau summer monsoon northward extent revealed by measurements of water stable isotopes
1
2001
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
Oxygen isotope ratios of sedimentary biogenic silica reflect the European transcontinental climate gradient
2
2008
... 近年来,Chen等(2016)建立了青藏高原中南部区域上多个湖泊中现生萝卜螺属壳体地球化学与其宿生水体水化学之间的关系,即萝卜螺属壳体δ18Oshell的变化主要反映了湖水氧同位素(δ18Owater)的变化,例如湖水蒸发或者降水δ18O的变化(Tyler et al, 2008);萝卜螺属壳体δ13Cshell取决于湖水的溶解无机碳(DIC)的δ13CDIC,并初步探讨了萝卜螺属壳体—水体δ18O分馏系数(变化范围在1.031~1.040)及其温度依赖性(较低).在此基础上,Chen等(2017)利用纳木错周边的高湖面沉积物中的萝卜螺属壳体化石的δ18Oshell和δ13Cshell结合高湖面沉积物的理化性质探讨了纳木错中晚全新世高湖面的变化机制. ...
... 在青藏高原,萝卜螺壳体δ18Oshell主要受宿生水体的δ18Owater控制(Chen et al, 2016).水体δ18Owater主要受降水的δ18Owater控制(Tyler et al, 2008),其他影响因素有:水温、蒸发、水体大小、湿度以及居留时间δ18Owater影响因素众多,水汽来源也较为复杂(Tian et al, 2001; Hu et al, 2017).大多数湖泊水体的δ18Owater受到区域降水和入湖水流的δ18Owater影响.但强烈的蒸发作用和居留时间使得不同水体δ18Owater产生较大的差异,进而使封闭湖泊中萝卜螺δ18Oshell比开放湖泊中萝卜螺δ18Oshell偏正(Taft et al, 2013).因此,鉴于萝卜螺属壳体δ18Oshell与其宿生湖泊水文状态存在一定的关系(Hu et al, 2017; 陈锋等, 2017)(图4),萝卜螺属壳体δ18Oshell可用于判断水体的水环境(封闭或者开放).河流、池塘、过水湖、半封闭湖和封闭湖中现生壳体δ18Oshell值的变化范围分别为:-16.2‰~ -15.5‰(Hu et al, 2017)、-14.6‰~-14.5‰(Hu et al, 2017)、-12.1‰~ -10.9‰(Chen et al, 2016)、-5.9‰~ -5.8‰(Chen et al, 2016)和-7.3‰~ -0.9‰(Chen et al, 2016).可见,萝卜螺属壳体的δ18Oshell值随湖泊水文状态开放程度的提高而逐渐降低.基于此,陈锋等(2017)应用Chen等(2016)建立的相关关系并结合纳木错高湖面沉积物中萝卜螺属的壳体化石,定量重建了该湖中晚全新世古湖水的水化学特征;并利用萝卜螺属壳体化石δ18Oshell,结合古湖岸线地貌证据,讨论了古湖水水文状态,认为萝卜螺属化石壳体的δ18Oshell值指示,纳木错在约距今约4.4-2.2 ka的高湖面期间可能处于一种间歇性外流状态. ...
Freshwater biogeography and limnological evolution of the Tibetan Plateau-Insights from a plateau-wide distributed gastropod taxon (Radix spp.)
5
2011
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... ).近年来,随着分子生物学的发展,学者们开始利用DNA技术对现生萝卜螺属进行分类(von Oheimb et al, 2011; 王瑞爱等, 2013),使得分类结果的可靠性更高,进而有利于合理评价软体动物壳体同位素和元素种内和种间变化.一般地,利用软体动物壳体进行同位素和元素序列分析时,首先应尽量挑选相同属种的样品,其次为相同属不同种的样品,这样壳体同位素和元素才能够更好地指示古气候或古环境变化(宋磊等, 2013). ...
... 萝卜螺属倾向于栖息在地势平坦、平静的浅水地带,如湖水、河水的回水区、湿地等(Glöer, 2002).在青藏高原的湖泊中,它们倾向于栖息在平静的浅水地带,多集中在有水草的近岸地带,在0~1.5 m的水深范围内很常见(von Oheimb et al, 2011; Taft et al, 2012, 2013; Chen et al, 2016).根据Mischke、Almogi-Labin等(2010)在冬给措纳湖的调查,获得萝卜螺属壳体的最大水深是47.9 m,然而萝卜螺属壳体更高频的收集为水深小于25 m的地带.另外,Mischke、Bößneck等(2010)在卡拉库里湖(Lake Karakul)的调查发现,活体萝卜螺属的种的分布水深在5.7~22.2 m.目前,基于萝卜螺属的生境推断古环境信息已经有了新的进展,例如Hu等(2017)利用青藏高原雅鲁藏布江古堰塞湖沉积物中萝卜螺属的习性及其壳体化石的δ18Oshell,推断出古堰塞湖的回水末端位于日喀则附近;并进一步结合高精度 GPS 测量,证实古堰塞湖湖面海拔高度大约为 3811 m. ...
Stable isotopes in fossil mammals, fish and shells from Kunlun Pass Basin, Tibetan Plateau: Paleo-climatic and paleo-elevation implications
1
2008
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
Chemical and isotopic (O, H, C) composition of surface waters in the catchment of Lake Donggi Cona (NW China) and implications for paleoenvironmental reconstructions
2
2016
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
A Holocene molluscan succession from floodplain sediments of the upper Lena River (Lake Baikal region), Siberia
2
2008
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
Elemental composition of carbonate skeletons, minerals, and sediments
1
1967
... 前人研究已表明,软体动物壳体的Sr/Ca比值与其生活水域的水体的温度(Dodd, 1965; Surge et al, 2006)或者盐度(Wolf et al, 1967; Eisma et al, 1976)存在显著的关系.Chen等(2016)对青藏高原中南部湖泊中现生萝卜螺属壳体元素与其宿生水体元素之间的关系进行了探讨,结果表明:萝卜螺属壳体的Sr/Cashell与其生活水体的Sr/Cawater呈显著的线性正相关(Sr/Cashell=0.118×Sr/Cawater+0.0008; R2=0.78);Sr/Cashell与湖水的电导率(EC)也呈显著的线性正相关(Sr/Cashell=9×10-7×EC+0.0001; R2=0.71);Sr/Cashell与其生活水体的温度和pH值均没有明显的关系;现生萝卜螺属壳体Sr的分配系数的变化范围在0.167~0.867之间,平均值是0.317,这一分配系数与以往的研究结果相接近.基于萝卜螺属壳体与宿生水体已建立的地球化学关系,陈锋等(2017)结合纳木错高湖面沉积物中萝卜螺属壳体化石定量重建了该湖4.4-2.2 ka期间湖水的古Sr/CaPalaeo-water和古电导率. ...
Morphodynamics and lake level variations at Paiku Co, southern Tibetan Plateau, China
2
2015
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
... 前人对萝卜螺属的研究主要集中在其生态(王泽长, 1963; 李朝品, 1998; 李宽意等, 2008; 刘海平等, 2014)、生物学(von Oheimb et al, 2011; 王瑞爱等, 2013)、寄生虫(Jouet et al, 2010; Huňová et al, 2012)、地理分布及经济意义(刘月英等, 1979)等方面的研究.同时开始涉及其壳体的年代学(李炳元等, 1982; 朱大岗等, 2003; Mischke et al, 2013; Wünnemann et al, 2015)和地层学(White et al, 2008; 李保生等, 2010; 韩建恩等, 2012)方面的研究.近年来,学者们才逐步开展利用萝卜螺属壳体进行环境重建方面的研究(Mischke et al, 2002; Taft et al, 2014; Weynell et al, 2016; Chen et al, 2017; 陈锋等, 2017). ...
Diatom-based conductivity and water-level inference models from eastern Tibetan (Qinghai-Xizang) Plateau lakes
1
2003
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
The life cycles of six species of freshwater molluscs in the Worcester-Birmingham Canal
2
1975
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
Slowing extrusion tectonics: Lowered estimate of post-Early Miocene slip rate for the Altyn Tagh fault
1
2004
... 值得注意的是,在全球众多现代湖泊及河流中发现有大型水生腹足类生物萝卜螺属(Radix)生存(刘月英等, 1979; Jokinen, 1992; von Oheimb et al, 2011; 刘海平等, 2014; Chen et al, 2016)(图1);同时其壳体化石亦常见于第三纪以来的湖泊沉积、河湖相沉积及高湖面沉积物中(Yue et al, 2004; Wang et al, 2008; 韩建恩等, 2012; Huntington et al, 2015; Weynell et al, 2016)(图2).特别是,萝卜螺属壳体化石广泛地分布于青藏高原全新世的湖相沉积物及湖岸阶地中(李炳元等, 1982; Hudson et al, 2015; Mischke et al, 2015; Wünnemann et al, 2015; Chen et al, 2017).萝卜螺属碳酸盐壳体的矿物组成为文石;并且萝卜螺属全新世时期的壳体化石的XRD衍射表明没有发生相变(陈锋, 2016).而萝卜螺属的壳体就可记录年、季甚至周内的湖泊水化学、水文和气候信息(Taft et al, 2012; Taft et al, 2013; Taft et al, 2014). ...
A chironomid-based salinity inference model from lakes on the Tibetan Plateau
1
2007
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
Biological and ecological features of saline lakes in northern Tibet, China
1
2005
... 萝卜螺属是淡水中常见的螺类,寿命约为1年(Young, 1975),其壳体增长速率夏季约为2.41~2.86 mm/月;冬季约为0.35 mm/月(Gaten, 1986).萝卜螺属能够记录高分辨率的环境信息正是得益于其较慢的生长速率(Taft et al, 2012).萝卜螺属个体越大,生活时间越长;其活动时间约为3-11月份,其中7-10月为高峰期,孳生密度很高(李朝品, 1998).因而,要想捕获现生萝卜螺属进行研究,最好在7-8月份进行.值得注意的是,除了地理分布广以外,萝卜螺属对水的酸碱度适应性也较强,pH值在5.8~9.9范围内皆可孳生,最适的pH值范围是7.0~9.6(Økland, 1990).尽管Zhao等(2005)在西藏北部盐度为20 g/L 的超咸水体(Tai Co)中发现了萝卜螺属的活动,但它们更喜欢生活在淡水或者低到中盐度的水体中(Taft et al, 2012).因此,基于这一特性,在古环境研究中,萝卜螺属等生物化石的发现,本身就说明了重要的环境信息.例如,位于青藏高原北部的柴达木盆地(36. 9°N; 95. 3°E)察尔汗盐湖东侧的贝壳层沉积中的腹足类、瓣鳃类和介形类古生物化石记录表明,当时该湖为淡水—微咸水,说明至少在38-28 ka BP之间,察尔汗古湖湖泊范围较大、湖面较高,为水质较淡的大湖(李炳元, 2000).Chen等(2017)对纳木错高湖面沉积物进行了多指标研究,认为在大约4.4-2.2 ka 期间,纳木错比现今湖泊范围更大、湖面更高,为水质更淡的大湖,并且这些沉积物中广泛分布的萝卜螺属壳体化石进一步支持了这一结论. ...
Ostracod-based environmental reconstruction over the last 8400 years of Nam Co Lake on the Tibetan Plateau
1
2010
... 在全球变化背景下,湖泊水文、水化学及其反映的气候变化得到了广泛关注(张灿等, 2015; 刘翀等, 2017; 傅伯杰, 2018; 杨大文等, 2018).在定量重建湖泊古水文、古水化学和古气候的研究中,湖泊沉积物中的化石生物指标发挥了极为重要的作用(沈吉等, 2010),如孢粉组合(Lü et al, 2011; Chen et al, 2015)、硅藻组合(Yang et al, 2003; 羊向东等, 2003)、摇蚊组合(张恩楼, 2006; Zhang et al, 2007)、介形类组合(Mischke, Almogi-Labin et al, 2010; Mischke, Bößneck et al, 2010; Zhu et al, 2010)、生物标志化合物指标(Liu et al, 2006; Hou et al, 2016)等.然而,在高原地区,已报道的古水文、古水化学和古气候定量重建数据的时间分辨率一般是百年或者几十年尺度(Taft et al, 2012).目前,该地区还较缺少记录过去水文、水化学和气候变化信息的高分辨率(年、季,甚至周)的理想载体. ...
De Nederlandse zoetwatermollusken: Recente en fossiele weekdieren uit zoet en brak water
1
1998
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
Vergleichende Untersuchung der Di?ten von Lymnaea auricularia undL. peregra (Gastropoda: Basommatophora) im Zürichsee
1
1977
... 萝卜螺属属于少有的能栖息在极端寒冷环境的腹足类动物,White等(2008)发现其可在冰雪覆盖超过半年的西伯利亚中南部的Basovo湖(55°52′N, 105°47′E; 345 m)中生存.Gaten(1986)观察到,在冬季萝卜螺属为了避免湖冰的侵袭或影响,会从浅水区向深水区移动.因此,萝卜螺属可以在寒冷的水环境中存活(Glöer, 2002; Young, 1975).同时,这种习性在古环境研究中也得到了验证.Hu等(2017)在雅鲁藏布江中游河谷阶地及古堰塞湖的研究中证实了在青藏高原萝卜螺属度过了末次冰期的寒冷气候期.值得注意的是,海拔对于萝卜螺属的生存似乎影响不大,在超高海拔的湖泊普莫雍错(5030 m; 表1)和亚当错(海拔5060 m; 表1)中也能发现萝卜螺属,甚至在海拔超过6000 m的地方也有它们的分布(刘月英等, 1979).总之,在青藏高原,发现有萝卜螺属活动的湖泊,其海拔大都集中在3000~6000 m.Taft等(2012)认为,萝卜螺属生长的限制因子不是低温而是营养.萝卜螺属的食物来源主要有绿藻门、蓝藻细菌、硅藻、原生动物、杂草(Gittenberger et al, 1998).因此,在萝卜螺属野外调查过程中,要优先在水草等植物密集的地区勘查,因为它们每天的活动范围依赖于食物的供给(Knecht et al, 1977).另外,笔者野外观察到,在光照充足的中午,萝卜螺属偶尔会出露在近岸或者近岸的水生植物上. ...
 , 冯金良
, 冯金良


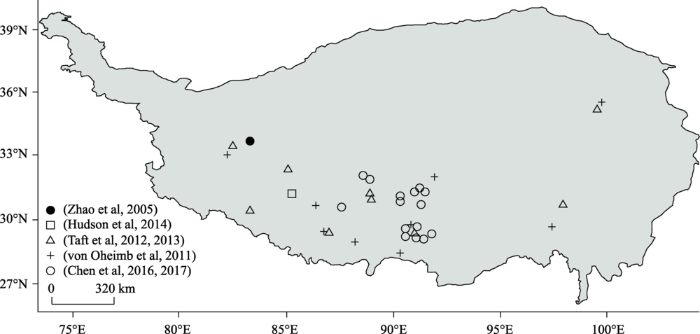
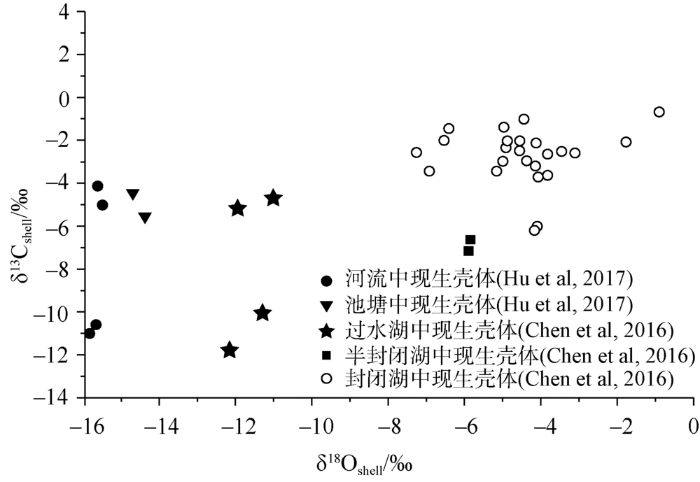
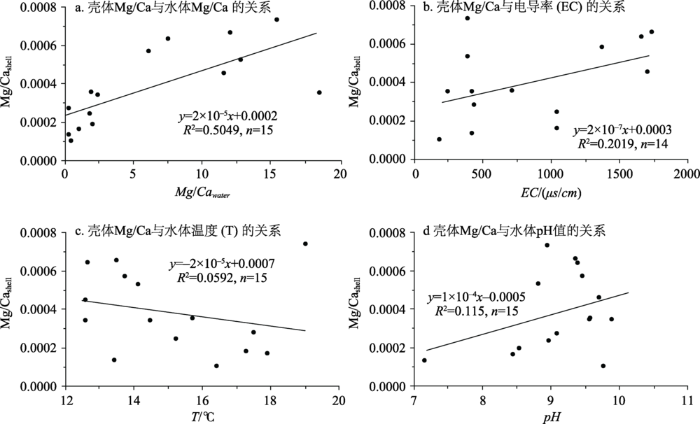
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}